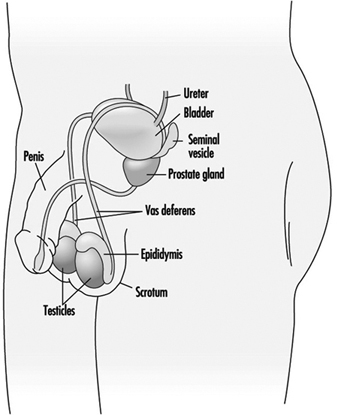
La espermatogénesis y la espermiogénesis son los procesos celulares que producen células sexuales masculinas maduras. Estos procesos tienen lugar dentro de los túbulos seminíferos de los testículos del macho sexualmente maduro, como se muestra en la Figura 1. Los túbulos seminíferos humanos tienen de 30 a 70 cm de largo y de 150 a 300 mm de diámetro (Zaneveld 1978). Las espermatogonias (células madre) se sitúan a lo largo de la membrana basal de los túbulos seminíferos y son las células básicas para la producción de espermatozoides.
Figura 1. El sistema reproductor masculino
Los espermatozoides maduran a través de una serie de divisiones celulares en las que las espermatogonias proliferan y se convierten en espermatocitos primarios. Los espermatocitos primarios en reposo migran a través de uniones estrechas formadas por las células de Sertoli hacia el lado luminal de esta barrera testicular. Cuando los espermatocitos alcanzan la barrera de la membrana en los testículos, la síntesis de ADN, el material genético en el núcleo de la célula, está esencialmente completa. Cuando los espermatocitos primarios se encuentran realmente con la luz del túbulo seminífero, estos experimentan un tipo especial de división celular que ocurre solo en las células germinales y se conoce como meiosis. La división celular meiótica da como resultado la división de los pares de cromosomas en el núcleo, de modo que cada célula germinal resultante contiene solo una copia de cada hebra cromosómica en lugar de un par coincidente.
Durante la meiosis, los cromosomas cambian de forma al condensarse y volverse filamentosos. En un momento dado, la membrana nuclear que los rodea se rompe y los husos microtubulares se unen a los pares cromosómicos, lo que hace que se separen. Esto completa la primera división meiótica y se forman dos espermatocitos secundarios haploides. Luego, los espermatocitos secundarios se someten a una segunda división meiótica para formar un número igual de espermátidas portadoras de cromosomas X e Y.
La transformación morfológica de las espermátidas en espermatozoides se denomina espermiogénesis. Cuando se completa la espermiogénesis, la célula de Sertoli libera cada espermatozoide en la luz de los túbulos seminíferos mediante un proceso denominado espermiación. Los espermatozoides migran a lo largo del túbulo hasta la rete testis y hacia la cabeza del epidídimo. Los espermatozoides que salen de los túbulos seminíferos son inmaduros: incapaces de fertilizar un óvulo e incapaces de nadar. Los espermatozoides liberados en la luz del túbulo seminífero están suspendidos en un líquido producido principalmente por las células de Sertoli. Los espermatozoides concentrados suspendidos dentro de este fluido fluyen continuamente desde los túbulos seminíferos, a través de ligeros cambios en el medio iónico dentro de la rete testis, a través de los conductos eferentes y hacia el epidídimo. El epidídimo es un solo tubo muy enrollado (de cinco a seis metros de largo) en el que los espermatozoides pasan de 12 a 21 días.
Dentro del epidídimo, los espermatozoides adquieren progresivamente motilidad y capacidad fecundante. Esto puede deberse a la naturaleza cambiante del fluido en suspensión en el epidídimo. Es decir, a medida que las células maduran, el epidídimo absorbe componentes del líquido, incluidas las secreciones de las células de Sertoli (p. ej., proteína fijadora de andrógenos), aumentando así la concentración de espermatozoides. El epidídimo también contribuye con sus propias secreciones al líquido de suspensión, incluidas las sustancias químicas glicerilfosforilcolina (GPC) y carnitina.
La morfología del esperma continúa transformándose en el epidídimo. La gotita citoplasmática se desprende y el núcleo del espermatozoide se condensa aún más. Si bien el epidídimo es el principal reservorio de almacenamiento de esperma hasta la eyaculación, alrededor del 30% de los espermatozoides en una eyaculación se han almacenado en los conductos deferentes. La eyaculación frecuente acelera el paso de los espermatozoides a través del epidídimo y puede aumentar la cantidad de espermatozoides inmaduros (estériles) en el eyaculado (Zaneveld 1978).
Eyaculación
Una vez dentro de los conductos deferentes, los espermatozoides son transportados por las contracciones musculares de la eyaculación más que por el flujo de líquido. Durante la eyaculación, los fluidos son expulsados a la fuerza de las glándulas sexuales accesorias dando lugar al plasma seminal. Estas glándulas no expulsan sus secreciones al mismo tiempo. Más bien, la glándula bulbouretral (de Cowper) primero expulsa un líquido transparente, seguido de las secreciones prostáticas, los líquidos concentrados en espermatozoides de los epidídimos y la ampolla de los conductos deferentes y, finalmente, la fracción más grande, principalmente de las vesículas seminales. Así, el plasma seminal no es un fluido homogéneo.
Acciones tóxicas sobre la espermatogénesis y la espermiogénesis
Los tóxicos pueden interrumpir la espermatogénesis en varios puntos. Los más dañinos, debido a su irreversibilidad, son los tóxicos que matan o alteran genéticamente (más allá de los mecanismos de reparación) las espermatogonias o las células de Sertoli. Los estudios en animales han sido útiles para determinar la etapa en la que un tóxico ataca el proceso espermatogénico. Estos estudios emplean exposición a corto plazo a un tóxico antes del muestreo para determinar el efecto. Al conocer la duración de cada etapa espermatogénica, se puede extrapolar para estimar la etapa afectada.
El análisis bioquímico del plasma seminal proporciona información sobre la función de las glándulas sexuales accesorias. Los productos químicos que son secretados principalmente por cada una de las glándulas sexuales accesorias se seleccionan típicamente para que sirvan como marcador para cada glándula respectiva. Por ejemplo, el epidídimo está representado por GPC, las vesículas seminales por fructosa y la glándula prostática por zinc. Tenga en cuenta que este tipo de análisis proporciona solo información general sobre la función glandular y poca o ninguna información sobre los demás constituyentes secretores. La medición del pH y la osmolalidad del semen proporciona información general adicional sobre la naturaleza del plasma seminal.
El plasma seminal puede analizarse para detectar la presencia de un tóxico o su metabolito. Se han detectado metales pesados en el plasma seminal utilizando espectrofotometría de absorción atómica, mientras que los hidrocarburos halogenados se han medido en fluido seminal mediante cromatografía de gases después de la extracción o filtración con limitación de proteínas (Stachel et al. 1989; Zikarge 1986).
La viabilidad y motilidad de los espermatozoides en el plasma seminal es típicamente un reflejo de la calidad del plasma seminal. Las alteraciones en la viabilidad de los espermatozoides, medida por exclusión de tinción o por hinchazón hipoosmótica, o alteraciones en los parámetros de motilidad de los espermatozoides sugerirían efectos tóxicos postesticulares.
Los análisis de semen también pueden indicar si la producción de espermatozoides se ha visto afectada por un tóxico. El recuento de espermatozoides y la morfología de los espermatozoides proporcionan índices de la integridad de la espermatogénesis y la espermiogénesis. Por lo tanto, el número de espermatozoides en el eyaculado está directamente relacionado con el número de células germinales por gramo de testículo (Zukerman et al. 1978), mientras que la morfología anormal es probablemente el resultado de una espermiogénesis anormal. Los espermatozoides muertos o los espermatozoides inmóviles a menudo reflejan los efectos de los eventos postesticulares. Por lo tanto, el tipo o momento de un efecto tóxico puede indicar el objetivo del tóxico. Por ejemplo, la exposición de ratas macho al 2-metoxietanol resultó en una reducción de la fertilidad después de cuatro semanas (Chapin et al. 1985). Esta evidencia, corroborada por el examen histológico, indica que el objetivo de la toxicidad es el espermatocito (Chapin et al. 1984). Si bien no es ético exponer intencionalmente a los seres humanos a posibles tóxicos para la reproducción, los análisis de semen de eyaculados en serie de hombres expuestos inadvertidamente durante un breve período de tiempo a posibles tóxicos pueden proporcionar información útil similar.
La exposición ocupacional al 1,2-dibromocloropropano (DBCP) redujo la concentración de esperma en los eyaculados de una mediana de 79 millones de células/ml en hombres no expuestos a 46 millones de células/ml en trabajadores expuestos (Whorton et al. 1979). Al retirar a los trabajadores de la exposición, aquellos con recuentos reducidos de espermatozoides experimentaron una recuperación parcial, mientras que los hombres que habían sido azoospérmicos permanecieron estériles. La biopsia testicular reveló que el objetivo de DBCP eran las espermatogonias. Esto corrobora la gravedad del efecto cuando las células madre son el objetivo de los tóxicos. No hubo indicios de que la exposición de los hombres a DBCP estuviera asociada con un resultado adverso del embarazo (Potashnik y Abeliovich 1985). Otro ejemplo de un tóxico dirigido a la espermatogénesis/espermiogénesis fue el estudio de trabajadores expuestos al dibromuro de etileno (EDB). Tenían más espermatozoides con cabezas cónicas y menos espermatozoides por eyaculado que los controles (Ratcliffe et al. 1987).
El daño genético es difícil de detectar en el esperma humano. Varios estudios en animales que utilizan el ensayo letal dominante (Ehling et al. 1978) indican que la exposición paterna puede producir un resultado adverso en el embarazo. Los estudios epidemiológicos de grandes poblaciones han demostrado una mayor frecuencia de abortos espontáneos en mujeres cuyos maridos trabajaban como mecánicos de vehículos motorizados (McDonald et al. 1989). Dichos estudios indican la necesidad de métodos para detectar daño genético en el esperma humano. Estos métodos están siendo desarrollados por varios laboratorios. Estos métodos incluyen sondas de ADN para discernir mutaciones genéticas (Hecht 1987), cariotipo de cromosomas espermáticos (Martin 1983) y evaluación de la estabilidad del ADN mediante citometría de flujo (Evenson 1986).
Figura 2. Exposiciones asociadas positivamente con efectos adversos en la calidad del semen

La figura 2 enumera las exposiciones que se sabe que afectan la calidad del esperma y la tabla 1 proporciona un resumen de los resultados de los estudios epidemiológicos de los efectos paternos en los resultados reproductivos.
Tabla 1. Estudios epidemiológicos de los efectos paternos en el resultado del embarazo
| Referencia | Tipo de exposición u ocupación | Asociación con la exposición1 | Efecto |
| Estudios de población basados en registros | |||
| Lindbohm et al. 1984 | disolventes | – | Aborto espontáneo |
| Lindbohm et al. 1984 | Estación de servicio | + | Aborto espontáneo |
| Daniell y Vaughan 1988 | Disolventes orgánicos | – | Aborto espontáneo |
| McDonald et al. 1989 | La Mecánica | + | Aborto espontáneo |
| McDonald et al. 1989 | Procesamiento de alimentos | + | defectos de desarrollo |
| Lindbohm et al. 1991a | Óxido de etileno | + | Aborto espontáneo |
| Lindbohm et al. 1991a | refinería de petróleo | + | Aborto espontáneo |
| Lindbohm et al. 1991a | Impregnaciones de madera | + | Aborto espontáneo |
| Lindbohm et al. 1991a | Productos químicos de caucho | + | Aborto espontáneo |
| Olsen et al. 1991 | Metales | + | Riesgo de cáncer infantil |
| Olsen et al. 1991 | maquinistas | + | Riesgo de cáncer infantil |
| Olsen et al. 1991 | Smiths | + | Riesgo de cáncer infantil |
| Kristensen et al. 1993 | disolventes | + | Parto prematuro |
| Kristensen et al. 1993 | plomo y solventes | + | Parto prematuro |
| Kristensen et al. 1993 | Lidera | + | Muerte perinatal |
| Kristensen et al. 1993 | Lidera | + | Morbilidad del niño varón |
| Estudios de casos y controles | |||
| kucera 1968 | Industria de la impresión | (+) | Labio leporino |
| kucera 1968 | Pintar | (+) | Paladar hendido |
| Olsen 1983 | Pintar | + | Daño al sistema nervioso central |
| Olsen 1983 | disolventes | (+) | Daño al sistema nervioso central |
| Sever et al. 1988 | Radiación de bajo nivel | + | Defectos del tubo neural |
| Taskinen et al. 1989 | Disolventes orgánicos | + | Aborto espontáneo |
| Taskinen et al. 1989 | Hidrocarbonos aromáticos | + | Aborto espontáneo |
| Taskinen et al. 1989 | Dust | + | Aborto espontáneo |
| Garner et al. 1990 | La radiación | + | Leucemia infantil |
| Bonde 1992 | Soldadura | + | Tiempo hasta la concepción |
| Wilkins y sumideros 1990 | Agricultura | (+) | tumor cerebral infantil |
| Wilkins y sumideros 1990 | Construcción | (+) | tumor cerebral infantil |
| Wilkins y sumideros 1990 | Procesamiento de alimentos/tabaco | (+) | tumor cerebral infantil |
| Wilkins y sumideros 1990 | Metal | + | tumor cerebral infantil |
| Lindböhmn et al. 1991b | Lidera | (+) | Aborto espontáneo |
| Sallmen et al. 1992 | Lidera | (+) | Defectos congénitos |
| Veulemans et al. 1993 | Éter de etilenglicol | + | Espermiograma anormal |
| Chia y col. 1992 | Metales | + | Cadmio en semen |
1 – ninguna asociación significativa; (+) asociación marginalmente significativa; + asociación significativa.
Fuente: Adaptado de Taskinen 1993.
Sistema Neuroendocrino
El funcionamiento general del sistema reproductivo está controlado por el sistema nervioso y las hormonas producidas por las glándulas (el sistema endocrino). El eje neuroendocrino reproductivo del varón involucra principalmente el sistema nervioso central (SNC), la glándula pituitaria anterior y los testículos. Las entradas del SNC y de la periferia están integradas por el hipotálamo, que regula directamente la secreción de gonadotropinas por parte de la hipófisis anterior. Las gonadotropinas, a su vez, actúan principalmente sobre las células de Leydig dentro del intersticio y las células de Sertoli y germinales dentro de los túbulos seminíferos para regular la espermatogénesis y la producción de hormonas en los testículos.
Eje hipotálamo-pituitario
El hipotálamo secreta la neurohormona hormona liberadora de gonadotropina (GnRH) en la vasculatura porta hipofisaria para su transporte a la glándula pituitaria anterior. La secreción pulsátil de este decapéptido provoca la liberación concomitante de hormona luteinizante (LH) y, con menor sincronía y una quinta parte de la potencia, la liberación de hormona folículo estimulante (FSH) (Bardin 1986). Existe evidencia sustancial para apoyar la presencia de una hormona liberadora de FSH separada, aunque todavía no se ha aislado ninguna (Savy-Moore y Schwartz 1980; Culler y Negro-Vilar 1986). Estas hormonas son secretadas por la glándula pituitaria anterior. La LH actúa directamente sobre las células de Leydig para estimular la síntesis y liberación de testosterona, mientras que la FSH estimula la aromatización de testosterona a estradiol por parte de las células de Sertoli. La estimulación gonadotrópica provoca la liberación de estas hormonas esteroides en la vena espermática.
La secreción de gonadotropina es, a su vez, controlada por la testosterona y el estradiol a través de mecanismos de retroalimentación negativa. La testosterona actúa principalmente sobre el hipotálamo para regular la secreción de GnRH y, por lo tanto, reduce la frecuencia del pulso, principalmente, de la liberación de LH. El estradiol, por otro lado, actúa sobre la glándula pituitaria para reducir la magnitud de la liberación de gonadotropinas. A través de estos bucles de retroalimentación endocrina, la función testicular en general y la secreción de testosterona en particular se mantienen en un estado relativamente estable.
Eje pituitario-testicular
La LH y la FSH generalmente se consideran necesarias para la espermatogénesis normal. Presumiblemente, el efecto de la LH es secundario a la inducción de altas concentraciones intratesticulares de testosterona. Por lo tanto, la FSH de la hipófisis y la testosterona de las células de Leydig actúan sobre las células de Sertoli dentro del epitelio de los túbulos seminíferos para iniciar la espermatogénesis. La producción de espermatozoides persiste, aunque se reduce cuantitativamente, después de eliminar la LH (y presumiblemente las altas concentraciones intratesticulares de testosterona) o la FSH. La FSH es necesaria para iniciar la espermatogénesis en la pubertad y, en menor medida, para reiniciar la espermatogénesis detenida (Matsumoto 1989; Sharpe 1989).
El sinergismo hormonal que sirve para mantener la espermatogénesis puede implicar el reclutamiento por parte de la FSH de espermatogonias diferenciadas para entrar en la meiosis, mientras que la testosterona puede controlar etapas posteriores específicas de la espermatogénesis. La FSH y la testosterona también pueden actuar sobre las células de Sertoli para estimular la producción de uno o más factores paracrinos que pueden afectar el número de células de Leydig y la producción de testosterona por estas células (Sharpe 1989). La FSH y la testosterona estimulan la síntesis de proteínas por parte de las células de Sertoli, incluida la síntesis de proteína fijadora de andrógenos (ABP), mientras que la FSH sola estimula la síntesis de aromatasa e inhibina. La ABP se secreta principalmente en el líquido de los túbulos seminíferos y se transporta a la porción proximal de la cabeza del epidídimo, posiblemente sirviendo como transportador local de andrógenos (Bardin 1986). La aromatasa cataliza la conversión de testosterona en estradiol en las células de Sertoli y en otros tejidos periféricos.
La inhibina es una glicoproteína que consta de dos subunidades diferentes unidas por disulfuro, a y b. Aunque la inhibina inhibe preferentemente la liberación de FSH, también puede atenuar la liberación de LH en presencia de estimulación con GnRH (Kotsugi et al. 1988). FSH y LH estimulan la liberación de inhibina con una potencia aproximadamente igual (McLachlan et al. 1988). Curiosamente, la inhibina se secreta en la sangre de la vena espermática en forma de pulsos que son sincrónicos con los de la testosterona (Winters 1990). Esto probablemente no refleja las acciones directas de la LH o la testosterona sobre la actividad de las células de Sertoli, sino más bien los efectos de otros productos de las células de Leydig secretados en los espacios intersticiales o en la circulación.
La prolactina, que también es secretada por la glándula pituitaria anterior, actúa sinérgicamente con la LH y la testosterona para promover la función reproductiva masculina. La prolactina se une a receptores específicos en la célula de Leydig y aumenta la cantidad de complejo receptor de andrógenos dentro del núcleo de los tejidos sensibles a los andrógenos (Baker et al. 1977). La hiperprolactinemia se asocia con reducciones del tamaño de los testículos y la próstata, el volumen del semen y las concentraciones circulantes de LH y testosterona (Segal et al. 1979). La hiperprolactinemia también se ha asociado con impotencia, aparentemente independiente de la alteración de la secreción de testosterona (Thorner et al. 1977).
Si se miden metabolitos de hormonas esteroides en la orina, se debe considerar la posibilidad de que la exposición que se está estudiando pueda alterar el metabolismo de los metabolitos excretados. Esto es especialmente pertinente ya que la mayoría de los metabolitos se forman en el hígado, un objetivo de muchas sustancias tóxicas. El plomo, por ejemplo, redujo la cantidad de esteroides sulfatados que se excretaban en la orina (Apostoli et al. 1989). Los niveles sanguíneos de ambas gonadotropinas se elevan durante el sueño cuando el varón entra en la pubertad, mientras que los niveles de testosterona mantienen este patrón diurno durante la edad adulta en los hombres (Plant 1988). Por lo tanto, las muestras de sangre, orina o saliva deben recolectarse aproximadamente a la misma hora del día para evitar variaciones debidas a los patrones secretores diurnos.
Los efectos evidentes de la exposición tóxica dirigida al sistema neuroendocrino reproductivo tienen más probabilidades de revelarse a través de manifestaciones biológicas alteradas de los andrógenos. Las manifestaciones significativamente reguladas por andrógenos en el hombre adulto que pueden detectarse durante un examen físico básico incluyen: (1) retención de nitrógeno y desarrollo muscular; (2) mantenimiento de los genitales externos y órganos sexuales accesorios; (3) mantenimiento de la laringe agrandada y cuerdas vocales engrosadas que causan la voz masculina; (4) crecimiento de barba, vello axilar y púbico y recesión del vello temporal y calvicie; (5) libido y rendimiento sexual; (6) proteínas específicas de órganos en tejidos (p. ej., hígado, riñones, glándulas salivales); y (7) conducta agresiva (Bardin 1986). Las modificaciones en cualquiera de estos rasgos pueden indicar que la producción de andrógenos se ha visto afectada.
Ejemplos de efectos tóxicos
El plomo es un ejemplo clásico de un tóxico que afecta directamente al sistema neuroendocrino. Las concentraciones séricas de LH se elevaron en hombres expuestos al plomo durante menos de un año. Este efecto no progresó en hombres expuestos durante más de cinco años. Los niveles séricos de FSH no se vieron afectados. Por otro lado, los niveles séricos de ABP se elevaron y los de testosterona total se redujeron en hombres expuestos al plomo durante más de cinco años. Los niveles séricos de testosterona libre se redujeron significativamente después de la exposición al plomo durante tres a cinco años (Rodamilans et al. 1988). Por el contrario, las concentraciones séricas de LH, FSH, testosterona total, prolactina y 17-cetosteroides neutros totales no se alteraron en los trabajadores con niveles circulantes de plomo más bajos, aunque se alteró la frecuencia de distribución del conteo de espermatozoides (Assennato et al. 1986). .
La exposición de los pintores de astilleros al 2-etoxietanol también redujo el conteo de espermatozoides sin un cambio simultáneo en las concentraciones séricas de LH, FSH o testosterona (Welch et al. 1988). Por lo tanto, los tóxicos pueden afectar la producción de hormonas y las medidas de esperma de forma independiente.
Los trabajadores masculinos involucrados en la fabricación del nematocida DBCP experimentaron niveles séricos elevados de LH y FSH y reducción del recuento de espermatozoides y la fertilidad. Aparentemente, estos efectos son secuelas de las acciones del DBCP sobre las células de Leydig para alterar la producción o la acción de los andrógenos (Mattison et al. 1990).
Varios compuestos pueden ejercer toxicidad en virtud de la similitud estructural con las hormonas esteroides reproductivas. Por lo tanto, al unirse al receptor endocrino respectivo, los tóxicos pueden actuar como agonistas o antagonistas para alterar las respuestas biológicas. La clordecona (Kepone), un insecticida que se une a los receptores de estrógeno, redujo el conteo y la motilidad de los espermatozoides, detuvo la maduración de los espermatozoides y redujo la libido. Si bien es tentador sugerir que estos efectos se deben a que la clordecona interfiere con las acciones de los estrógenos a nivel neuroendocrino o testicular, en estos estudios no se demostró que los niveles séricos de testosterona, LH y FSH se alteraran de manera similar a los efectos de la terapia con estradiol. . El DDT y sus metabolitos también exhiben propiedades esteroides y podría esperarse que alteren la función reproductiva masculina al interferir con las funciones de las hormonas esteroides. Los xenobióticos como los bifenilos policlorados, los bifenilos polibromados y los pesticidas organoclorados también pueden interferir con las funciones reproductivas masculinas al ejercer una actividad agonista/antagonista estrogénica (Mattison et al. 1990).
función sexual
La función sexual humana se refiere a las actividades integradas de los testículos y las glándulas sexuales secundarias, los sistemas de control endocrino y los componentes conductuales y psicológicos de la reproducción (libido) basados en el sistema nervioso central. La erección, la eyaculación y el orgasmo son tres eventos distintos, independientes, fisiológicos y psicodinámicos que normalmente ocurren simultáneamente en los hombres.
Hay pocos datos confiables disponibles sobre los efectos de la exposición ocupacional en la función sexual debido a los problemas descritos anteriormente. Se ha demostrado que las drogas afectan cada una de las tres etapas de la función sexual masculina (Fabro 1985), lo que indica la posibilidad de que las exposiciones ocupacionales ejerzan efectos similares. Los antidepresivos, los antagonistas de la testosterona y los estimulantes de la liberación de prolactina reducen eficazmente la libido en los hombres. Los fármacos antihipertensivos que actúan sobre el sistema nervioso simpático inducen impotencia en algunos hombres, pero sorprendentemente, priapismo en otros. La fenoxibenzamina, un antagonista adrenoceptivo, se ha utilizado clínicamente para bloquear la emisión seminal pero no el orgasmo (Shilon, Paz y Homonnai 1984). Los fármacos antidepresivos anticolinérgicos permiten la emisión seminal mientras bloquean la eyección seminal y el orgasmo, lo que da como resultado que el plasma seminal se filtre desde la uretra en lugar de ser expulsado.
Las drogas recreativas también afectan la función sexual (Fabro 1985). El etanol puede reducir la impotencia al tiempo que mejora la libido. La cocaína, la heroína y las altas dosis de cannabinoides reducen la libido. Los opiáceos también retrasan o deterioran la eyaculación.
La amplia y variada gama de productos farmacéuticos que se ha demostrado que afectan el sistema reproductivo masculino respalda la idea de que los productos químicos que se encuentran en el lugar de trabajo también pueden ser tóxicos para la reproducción. Se necesitan métodos de investigación que sean confiables y prácticos para las condiciones de estudio de campo para evaluar esta importante área de toxicología reproductiva.