La toxicologie génétique, par définition, est l'étude de la façon dont les agents chimiques ou physiques affectent le processus complexe de l'hérédité. Les produits chimiques génotoxiques sont définis comme des composés capables de modifier le matériel héréditaire des cellules vivantes. La probabilité qu'un produit chimique particulier cause des dommages génétiques dépend inévitablement de plusieurs variables, dont le niveau d'exposition de l'organisme au produit chimique, la distribution et la rétention du produit chimique une fois qu'il pénètre dans l'organisme, l'efficacité des systèmes d'activation métabolique et/ou de détoxification dans tissus cibles et la réactivité du produit chimique ou de ses métabolites avec les macromolécules critiques dans les cellules. La probabilité qu'un dommage génétique cause une maladie dépend en fin de compte de la nature du dommage, de la capacité de la cellule à réparer ou à amplifier le dommage génétique, de la possibilité d'exprimer toute altération induite et de la capacité de l'organisme à reconnaître et à supprimer la multiplication des cellules aberrantes.
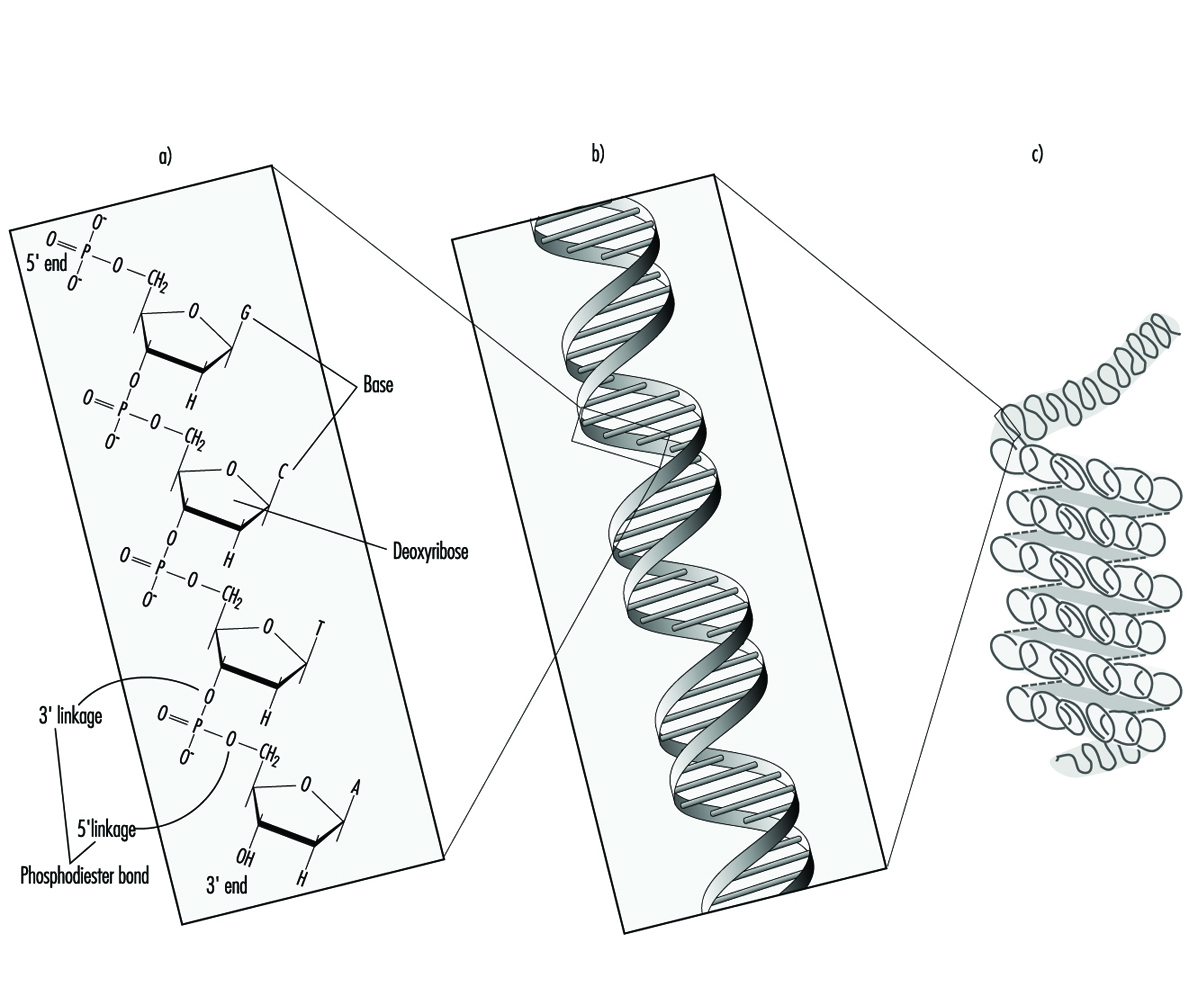
Dans les organismes supérieurs, l'information héréditaire est organisée en chromosomes. Les chromosomes sont constitués de brins étroitement condensés d'ADN associé à des protéines. Au sein d'un même chromosome, chaque molécule d'ADN existe sous la forme d'une paire de longues chaînes non ramifiées de sous-unités nucléotidiques reliées entre elles par des liaisons phosphodiester qui relient le carbone 5 d'un fragment désoxyribose au carbone 3 du suivant (figure 1). De plus, l'une des quatre bases nucléotidiques différentes (adénine, cytosine, guanine ou thymine) est attachée à chaque sous-unité désoxyribose comme des perles sur une ficelle. En trois dimensions, chaque paire de brins d'ADN forme une double hélice avec toutes les bases orientées vers l'intérieur de la spirale. Au sein de l'hélice, chaque base est associée à sa base complémentaire sur le brin d'ADN opposé ; la liaison hydrogène dicte un appariement fort et non covalent de l'adénine avec la thymine et de la guanine avec la cytosine (figure 1). Étant donné que la séquence des bases nucléotidiques est complémentaire sur toute la longueur de la molécule d'ADN duplex, les deux brins portent essentiellement la même information génétique. En effet, lors de la réplication de l'ADN chaque brin sert de matrice pour la production d'un nouveau brin partenaire.
Figure 1. L'organisation (a) primaire, (b) secondaire et (c) tertiaire de l'information héréditaire humaine
 À l'aide d'ARN et d'un ensemble de protéines différentes, la cellule déchiffre finalement les informations codées par la séquence linéaire de bases dans des régions spécifiques de l'ADN (gènes) et produit des protéines essentielles à la survie cellulaire de base ainsi qu'à la croissance et à la différenciation normales. Essentiellement, les nucléotides fonctionnent comme un alphabet biologique utilisé pour coder les acides aminés, les éléments constitutifs des protéines.
À l'aide d'ARN et d'un ensemble de protéines différentes, la cellule déchiffre finalement les informations codées par la séquence linéaire de bases dans des régions spécifiques de l'ADN (gènes) et produit des protéines essentielles à la survie cellulaire de base ainsi qu'à la croissance et à la différenciation normales. Essentiellement, les nucléotides fonctionnent comme un alphabet biologique utilisé pour coder les acides aminés, les éléments constitutifs des protéines.
Lorsque des nucléotides incorrects sont insérés ou que des nucléotides sont perdus, ou lorsque des nucléotides inutiles sont ajoutés pendant la synthèse de l'ADN, l'erreur est appelée mutation. Il a été estimé que moins d'une mutation se produit pour 109 nucléotides incorporés lors de la réplication normale des cellules. Bien que les mutations ne soient pas nécessairement nocives, les altérations entraînant l'inactivation ou la surexpression de gènes importants peuvent entraîner divers troubles, notamment le cancer, des maladies héréditaires, des anomalies du développement, l'infertilité et la mort embryonnaire ou périnatale. Très rarement, une mutation peut entraîner une amélioration de la survie ; de tels événements sont à la base de la sélection naturelle.
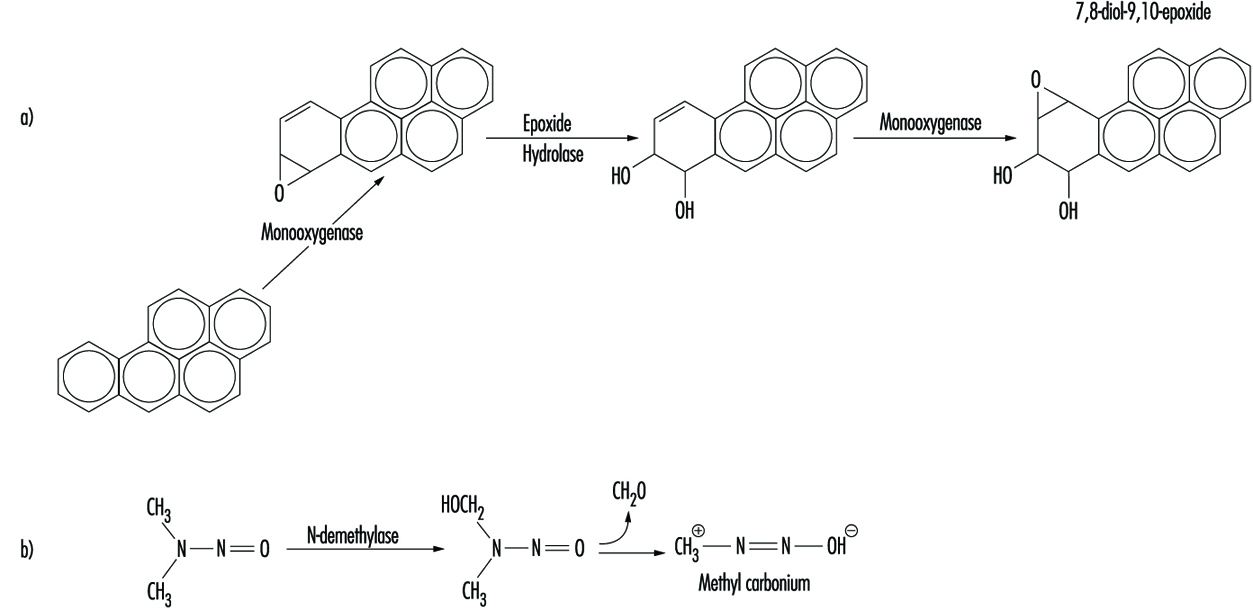
Bien que certains produits chimiques réagissent directement avec l'ADN, la plupart nécessitent une activation métabolique. Dans ce dernier cas, les intermédiaires électrophiles tels que les époxydes ou les ions carbonium sont finalement responsables de l'induction de lésions sur une variété de sites nucléophiles au sein du matériel génétique (figure 2). Dans d'autres cas, la génotoxicité est médiée par des sous-produits de l'interaction du composé avec des lipides intracellulaires, des protéines ou de l'oxygène.
Figure 2. Bioactivation de : a) benzo(a)pyrène ; et b) N-nitrosodiméthylamine
En raison de leur abondance relative dans les cellules, les protéines sont la cible la plus fréquente des interactions toxiques. Cependant, la modification de l'ADN est plus préoccupante en raison du rôle central de cette molécule dans la régulation de la croissance et de la différenciation à travers plusieurs générations de cellules.
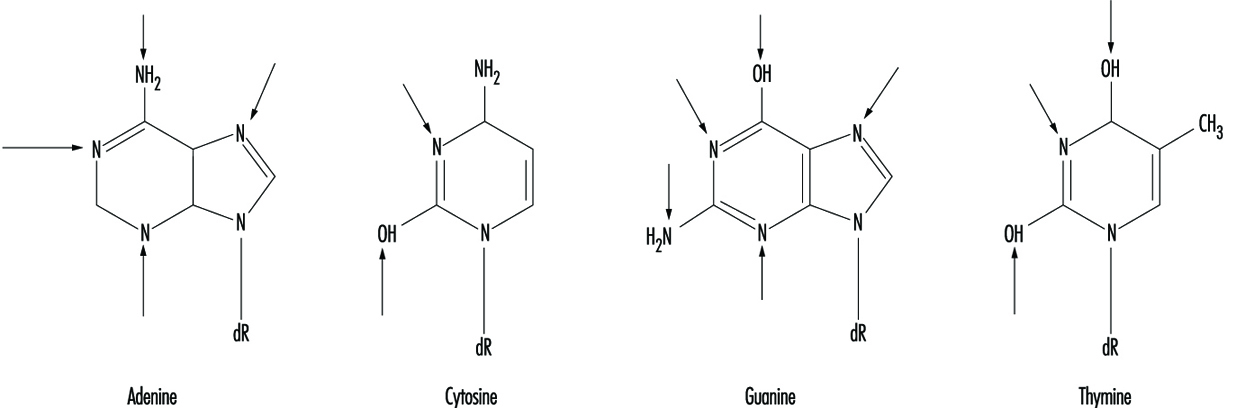
Au niveau moléculaire, les composés électrophiles ont tendance à attaquer l'oxygène et l'azote dans l'ADN. Les sites les plus susceptibles d'être modifiés sont illustrés à la figure 3. Bien que les oxygènes au sein des groupes phosphate dans le squelette de l'ADN soient également des cibles de modification chimique, on pense que les dommages aux bases sont biologiquement plus pertinents puisque ces groupes sont considérés comme le principal vecteur d'information. éléments de la molécule d'ADN.
Figure 3. Sites primaires de dommages à l'ADN induits chimiquement
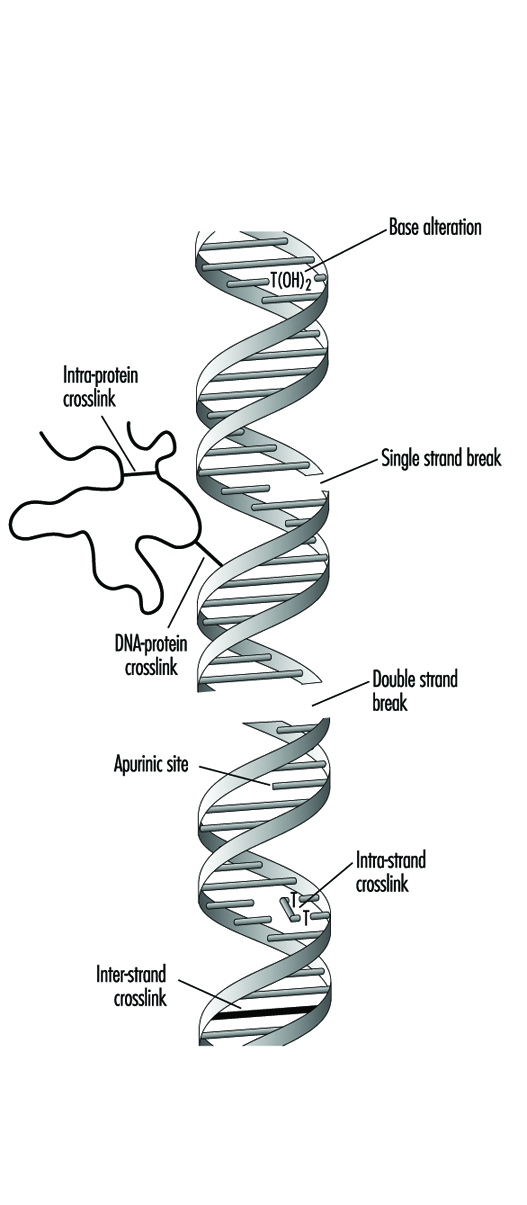
Les composés qui contiennent une fraction électrophile exercent généralement une génotoxicité en produisant des mono-adduits dans l'ADN. De même, les composés qui contiennent deux fractions réactives ou plus peuvent réagir avec deux centres nucléophiles différents et produire ainsi des réticulations intra- ou inter-moléculaires dans le matériel génétique (figure 4). Les réticulations interbrin ADN-ADN et ADN-protéine peuvent être particulièrement cytotoxiques car elles peuvent former des blocs complets pour la réplication de l'ADN. Pour des raisons évidentes, la mort d'une cellule élimine la possibilité qu'elle soit mutée ou transformée de façon néoplasique. Les agents génotoxiques peuvent également agir en induisant des cassures dans le squelette phosphodiester, ou entre les bases et les sucres (produisant des sites abasiques) dans l'ADN. De telles cassures peuvent être le résultat direct de la réactivité chimique au niveau du site endommagé ou peuvent se produire pendant la réparation de l'un des types de lésions de l'ADN susmentionnés.
Figure 4. Différents types de dommages au complexe protéine-ADN
Au cours des trente à quarante dernières années, diverses techniques ont été développées pour surveiller le type de dommages génétiques induits par divers produits chimiques. Ces tests sont décrits en détail ailleurs dans ce chapitre et Encyclopédie.
Une mauvaise réplication de « microlésions » telles que des mono-adduits, des sites abasiques ou des cassures simple brin peut finalement entraîner des substitutions de paires de bases de nucléotides, ou l'insertion ou la suppression de courts fragments de polynucléotides dans l'ADN chromosomique. En revanche, les «macrolésions», telles que les adduits volumineux, les réticulations ou les cassures double brin, peuvent déclencher le gain, la perte ou le réarrangement de morceaux de chromosomes relativement volumineux. Dans tous les cas, les conséquences peuvent être dévastatrices pour l'organisme puisque chacun de ces événements peut entraîner la mort cellulaire, la perte de fonction ou la transformation maligne des cellules. La manière exacte dont les dommages à l'ADN causent le cancer est en grande partie inconnue. On pense actuellement que le processus peut impliquer une activation inappropriée de proto-oncogènes tels que monc et de ras, et/ou l'inactivation de gènes suppresseurs de tumeurs récemment identifiés tels que p53. L'expression anormale de l'un ou l'autre type de gène abroge les mécanismes cellulaires normaux pour contrôler la prolifération et/ou la différenciation cellulaire.
La prépondérance des preuves expérimentales indique que le développement d'un cancer suite à une exposition à des composés électrophiles est un événement relativement rare. Cela peut s'expliquer, en partie, par la capacité intrinsèque de la cellule à reconnaître et à réparer l'ADN endommagé ou par l'incapacité des cellules dont l'ADN est endommagé à survivre. Pendant la réparation, la base endommagée, le nucléotide ou le court tronçon de nucléotides entourant le site endommagé est retiré et (en utilisant le brin opposé comme modèle) un nouveau morceau d'ADN est synthétisé et épissé en place. Pour être efficace, la réparation de l'ADN doit se produire avec une grande précision avant la division cellulaire, avant les opportunités de propagation de la mutation.
Des études cliniques ont montré que les personnes présentant des défauts héréditaires dans la capacité de réparer l'ADN endommagé développent fréquemment un cancer et/ou des anomalies du développement à un âge précoce (tableau 1). De tels exemples fournissent des preuves solides reliant l'accumulation de dommages à l'ADN à la maladie humaine. De même, les agents qui favorisent la prolifération cellulaire (tels que l'acétate de tétradécanoylphorbol) améliorent souvent la carcinogenèse. Pour ces composés, la probabilité accrue de transformation néoplasique peut être une conséquence directe d'une diminution du temps disponible pour que la cellule effectue une réparation adéquate de l'ADN.
Tableau 1. Troubles héréditaires prédisposés au cancer qui semblent impliquer des défauts de réparation de l'ADN
| Syndrome | Symptômes | Phénotype cellulaire |
| Ataxie télangiectasie | Détérioration neurologique Immunodéficience Incidence élevée de lymphome |
Hypersensibilité aux rayonnements ionisants et à certains agents alkylants. Réplication dérégulée de l'ADN endommagé (peut indiquer un temps raccourci pour la réparation de l'ADN) |
| Syndrome de Bloom | Anomalies du développement Lésions sur la peau exposée Incidence élevée de tumeurs du système immunitaire et du tractus gastro-intestinal |
Fréquence élevée des aberrations chromosomiques Ligature défectueuse des cassures associées à la réparation de l'ADN |
| L'anémie de Fanconi | Retard de croissance Incidence élevée de leucémie |
Hypersensibilité aux agents de réticulation Fréquence élevée des aberrations chromosomiques Réparation défectueuse des liaisons croisées dans l'ADN |
| Cancer du côlon héréditaire sans polypose | Forte incidence du cancer du côlon | Défaut dans la réparation des mésappariements d'ADN (lorsque l'insertion d'un mauvais nucléotide se produit pendant la réplication) |
| Xéroderma pigmentosum | Incidence élevée d'épithéliome sur les zones exposées de la peau Atteinte neurologique (dans de nombreux cas) |
Hypersensibilité aux rayons UV et à de nombreux cancérigènes chimiques Défauts de réparation par excision et/ou de réplication de l'ADN endommagé |
Les premières théories sur la façon dont les produits chimiques interagissent avec l'ADN remontent aux études menées lors du développement du gaz moutarde utilisé dans la guerre. Une meilleure compréhension est née des efforts visant à identifier des agents anticancéreux qui arrêteraient sélectivement la réplication des cellules tumorales à division rapide. L'inquiétude croissante du public concernant les dangers dans notre environnement a incité des recherches supplémentaires sur les mécanismes et les conséquences de l'interaction chimique avec le matériel génétique. Des exemples de divers types de produits chimiques qui exercent une génotoxicité sont présentés dans le tableau 2.
Tableau 2. Exemples de produits chimiques qui présentent une génotoxicité dans les cellules humaines
| Classe de produit chimique | Exemple | Source d'exposition | Lésion génotoxique probable |
| Aflatoxines | Aflatoxine B1 | Nourriture contaminée | Adduits volumineux à l'ADN |
| Amines aromatiques | 2-Acétylaminofluorène | Environnement | Adduits volumineux à l'ADN |
| Quinones d'aziridine | Mitomycine c | Chimiothérapie anticancéreuse | Mono-adduits, réticulations interbrins et cassures simple brin dans l'ADN. |
| Hydrocarbures chlorés | Chlorure de vinyle | Environnement | Mono-adduits dans l'ADN |
| Métaux et composés métalliques | Cisplatine | Chimiothérapie anticancéreuse | Crosslinks intra- et inter-brins dans l'ADN |
| Composés de nickel | Environnement | Mono-adduits et cassures simple brin dans l'ADN | |
| Moutardes à l'azote | Cyclophosphamide | Chimiothérapie anticancéreuse | Mono-adduits et réticulations interbrins dans l'ADN |
| Nitrosamines | N-Nitrosodiméthylamine | Nourriture contaminée | Mono-adduits dans l'ADN |
| Hydrocarbures aromatiques polycycliques | Benzo (a) pyrène | Environnement | Adduits volumineux à l'ADN |