Кожная чувствительность разделяет основные элементы всех основных органов чувств. Свойства внешнего мира, такие как цвет, звук или вибрация, воспринимаются специализированными окончаниями нервных клеток, называемыми сенсорными рецепторами, которые преобразуют внешние данные в нервные импульсы. Затем эти сигналы передаются в центральную нервную систему, где они становятся основой для интерпретации окружающего мира.
Полезно признать три основных момента, касающихся этих процессов. Во-первых, энергия и изменения энергетических уровней могут быть восприняты только органом чувств, способным обнаруживать конкретный тип рассматриваемой энергии. (Вот почему микроволны, рентгеновские лучи и ультрафиолетовые лучи опасны; мы не приспособлены для их обнаружения, так что даже на смертельных уровнях они не воспринимаются). нервная система ограничена восстановлением неполного изображения из сигналов, передаваемых ее сенсорными рецепторами. В-третьих, наши сенсорные системы предоставляют нам более точную информацию об изменениях в окружающей среде, чем о статических условиях. Мы хорошо оснащены сенсорными рецепторами, чувствительными, например, к мерцанию света или к малейшим колебаниям температуры, вызванным легким ветерком; мы менее приспособлены для получения информации, скажем, о постоянной температуре или о постоянном давлении на кожу.
Традиционно кожные ощущения делятся на две категории: кожные и глубокие. В то время как глубокая чувствительность зависит от рецепторов, расположенных в мышцах, сухожилиях, суставах и надкостнице (мембране, окружающей кости), кожная чувствительность, которую мы здесь рассматриваем, имеет дело с информацией, полученной рецепторами в коже: в частности, различными классами кожные рецепторы, расположенные на границе дермы и эпидермиса или рядом с ними.
Все сенсорные нервы, связывающие кожные рецепторы с центральной нервной системой, имеют примерно одинаковую структуру. Большое тело клетки находится в скоплении тел других нервных клеток, называемом ганглием, расположенном рядом со спинным мозгом и соединенном с ним узкой ветвью ствола клетки, называемой ее аксоном. Большинство нервных клеток или нейронов, которые берут начало в спинном мозге, посылают аксоны к костям, мышцам, суставам или, в случае кожной чувствительности, к коже. Подобно изолированному проводу, каждый аксон вдоль своего пути и на концах покрыт защитными слоями клеток, известных как шванновские клетки. Эти шванновские клетки производят вещество, известное как миелин, которое покрывает аксон как оболочку. Через определенные промежутки времени в миелиновой оболочке появляются крошечные разрывы, известные как узелки Ранвье. Наконец, на конце аксона находятся компоненты, специализирующиеся на получении и ретрансляции информации о внешней среде: сенсорные рецепторы (Mountcastle, 1974).
Различные классы кожных рецепторов, как и все сенсорные рецепторы, определяются двумя способами: их анатомическими структурами и типом электрических сигналов, которые они посылают по своим нервным волокнам. Определенно структурированные рецепторы обычно называют в честь их первооткрывателей. Относительно немногочисленные классы сенсорных рецепторов кожи можно разделить на три основные категории: механорецепторы, тепловые рецепторы и ноцицепторы.
Все эти рецепторы могут передавать информацию о конкретном раздражителе только после того, как они сначала закодировали его в виде электрохимического нейронного языка. Эти нейронные коды используют различные частоты и модели нервных импульсов, которые ученые только начали расшифровывать. Действительно, важная ветвь нейрофизиологических исследований полностью посвящена изучению сенсорных рецепторов и способов, которыми они переводят энергетические состояния в окружающей среде в нейронные коды. Как только коды сгенерированы, они передаются центрально по афферентным волокнам, нервным клеткам, которые обслуживают рецепторы, передавая сигналы в центральную нервную систему.
Сообщения, производимые рецепторами, можно разделить на основе ответа на непрерывную неизменную стимуляцию: медленно адаптирующиеся рецепторы посылают электрохимические импульсы в центральную нервную систему на время действия постоянного раздражителя, тогда как быстроадаптирующиеся рецепторы постепенно уменьшают свои разряды в присутствие устойчивого стимула до тех пор, пока они не достигнут низкого базового уровня или полностью не прекратятся, после чего перестанут информировать центральную нервную систему о продолжающемся присутствии стимула.
Совершенно разные ощущения боли, тепла, холода, давления и вибрации, таким образом, вызываются активностью различных классов сенсорных рецепторов и связанных с ними нервных волокон. Например, термины «трепетание» и «вибрация» используются для различения двух слегка различающихся вибрационных ощущений, кодируемых двумя разными классами чувствительных к вибрации рецепторов (Mountcastle et al., 1967). Каждая из трех важных категорий болевых ощущений, известных как колющая боль, жгучая боль и ноющая боль, связана с отдельным классом ноцицептивных афферентных волокон. Однако это не означает, что специфическое ощущение обязательно включает только один класс рецепторов; более чем один класс рецепторов может вносить вклад в данное ощущение, и фактически ощущения могут различаться в зависимости от относительного вклада разных классов рецепторов (Sinclair, 1981).
Предыдущее резюме основано на гипотезе специфичности кожной сенсорной функции, впервые сформулированной немецким врачом фон Фреем в 1906 году. в настоящее время сильно подтверждены фактическими доказательствами.
Рецепторы, реагирующие на постоянное кожное давление
В руке относительно крупные миелинизированные волокна (диаметром от 5 до 15 мм) выходят из подкожной нервной сети, называемой подпапиллярным нервным сплетением, и заканчиваются пучком нервных окончаний на стыке дермы и эпидермиса (рис. 1). В волосистой коже эти нервные окончания завершаются видимыми поверхностными структурами, известными как сенсорные купола; в гладкой или безволосой коже нервные окончания находятся у основания кожных гребней (например, образующих отпечатки пальцев). Там, в тактильном куполе, каждый кончик нервного волокна, или нейрит, окружен специализированной эпителиальной клеткой, известной как Ячейка Меркель (см. рисунки 2 и 3).
Рисунок 1. Схематическое изображение поперечного сечения кожи.

Рисунок 2. Сенсорный купол на каждом приподнятом участке кожи содержит от 30 до 70 клеток Меркеля.

Рисунок 3. При большем увеличении, доступном с помощью электронного микроскопа, видно, что клетка Меркеля, специализированная эпителиальная клетка, прикреплена к базальной мембране, которая отделяет эпидермис от дермы.

Комплекс нейритов клеток Меркеля преобразует механическую энергию в нервные импульсы. Хотя мало что известно о роли клетки или о ее механизме трансдукции, она была идентифицирована как медленно адаптирующийся рецептор. Это означает, что давление на сенсорный купол, содержащий клетки Меркеля, заставляет рецепторы генерировать нервные импульсы на время действия стимула. Эти импульсы возрастают по частоте пропорционально интенсивности раздражителя, тем самым информируя мозг о продолжительности и величине давления на кожу.
Как и клетка Меркеля, второй медленно адаптирующийся рецептор также служит коже, сигнализируя о величине и продолжительности устойчивого кожного давления. Видимый только в микроскоп, этот рецептор, известный как Рецептор Руффини, состоит из группы нейритов, выходящих из миелинового волокна и инкапсулированных клетками соединительной ткани. В структуре капсулы есть волокна, которые, по-видимому, передают локальные кожные искажения нейритам, которые, в свою очередь, передают сообщения по нервному тракту в центральную нервную систему. Давление на кожу вызывает устойчивый разряд нервных импульсов; как и в клетке Меркеля, частота нервных импульсов пропорциональна интенсивности раздражителя.
Несмотря на их сходство, между клетками Меркеля и рецепторами Руффини есть одно существенное различие. В то время как ощущения возникают при стимуляции рецепторов Руффини, стимуляция тактильных куполов, в которых находятся клетки Меркеля, не вызывает осознанных ощущений; сенсорный купол, таким образом, является загадочным рецептором, поскольку его фактическая роль в нервной функции остается неизвестной. Таким образом, рецепторы Руффини считаются единственными рецепторами, способными обеспечивать нервные сигналы, необходимые для сенсорного ощущения давления или постоянного прикосновения. Кроме того, было показано, что медленно адаптирующиеся рецепторы Руффини объясняют способность людей оценивать кожное давление по шкале интенсивности.
Рецепторы, реагирующие на вибрацию и движение кожи
В отличие от медленно адаптирующихся механорецепторов, быстро адаптирующиеся рецепторы молчат во время длительного вдавливания кожи. Однако они хорошо подходят для передачи сигналов вибрации и движения кожи. Отмечены две основные категории: на волосистой коже, связанные с отдельными волосками; и те, которые образуют корпускулярные окончания на голой или безволосой коже.
Рецепторы, обслуживающие волоски
Типичный волос окутан сетью нервных окончаний, отходящих от пяти до девяти крупных миелинизированных аксонов (рис. 4). У приматов эти окончания делятся на три категории: ланцетные окончания, веретенообразные окончания и папиллярные окончания. Все три быстро адаптируются, так что постоянное отклонение волос вызывает нервные импульсы только во время движения. Таким образом, эти рецепторы исключительно чувствительны к движущимся или вибрационным раздражителям, но дают мало или совсем не дают информации о давлении или постоянном прикосновении.
Рисунок 4. Стержни волосков являются площадкой для нервных окончаний, обнаруживающих движения.
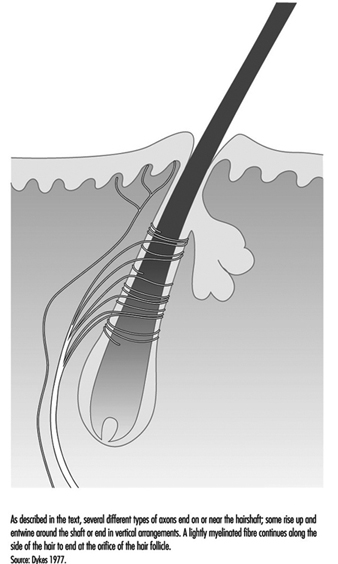
Ланцетные окончания возникают из сильно миелинизированных волокон, образующих сеть вокруг волос. Терминальные нейриты теряют свое обычное покрытие шванновскими клетками и проникают между клетками у основания волоса.
Веретенообразные окончания образованы окончаниями аксонов, окруженными шванновскими клетками. Терминалы поднимаются к наклонному стержню волоса и заканчиваются полукруглым скоплением чуть ниже сальной или сальной железы. Сосочковые окончания отличаются от веретенообразных окончаний тем, что они заканчиваются не на стержне волоса, а в виде свободных нервных окончаний вокруг устья волоса.
Предположительно, существуют функциональные различия между типами рецепторов, обнаруженных на волосах. Это можно вывести частично из структурных различий в том, как нервы заканчиваются на стержне волоса, и частично из различий в диаметре аксонов, поскольку аксоны разного диаметра соединяются с разными центральными релейными областями. Тем не менее, функции рецепторов волосистой кожи остаются предметом изучения.
Рецепторы гладкой кожи
Корреляция анатомической структуры рецептора с генерируемыми им нейронными сигналами наиболее выражена в крупных и легко управляемых рецепторах с корпускулярными или инкапсулированными окончаниями. Особенно хорошо изучены тельца пачининана и тельца Мейснера, которые, как и нервные окончания в волосах, о которых говорилось выше, передают ощущение вибрации.
Тельца Пачини достаточно велики, чтобы их можно было увидеть невооруженным глазом, что позволяет легко связать рецептор со специфическим нервным ответом. Расположенный в дерме, обычно вокруг сухожилий или суставов, он представляет собой луковицу размером 0.5 × 1.0 мм. Его обслуживает одно из самых крупных афферентных волокон в организме, имеющее диаметр от 8 до 13 мкм и проводящее со скоростью от 50 до 80 метров в секунду. Хорошо известна его анатомия, хорошо изученная как световой, так и электронной микроскопией.
Основным компонентом тельца является внешнее ядро, состоящее из клеточного материала, заключающего в себе заполненные жидкостью пространства. Само внешнее ядро окружено капсулой, пронизанной центральным каналом и капиллярной сетью. Через канал проходит одиночное миелинизированное нервное волокно диаметром от 7 до 11 мм, которое становится длинным немиелиновым нервным окончанием, проникающим глубоко в центр тельца. Конечный аксон эллиптический, с ветвящимися отростками.
Тельца Пачини — быстро адаптирующиеся рецепторы. Таким образом, при длительном давлении он производит импульс только в начале и в конце раздражителя. Он реагирует на высокочастотные вибрации (от 80 до 400 Гц) и наиболее чувствителен к вибрациям с частотой около 250 Гц. Часто эти рецепторы реагируют на вибрации, передаваемые по костям и сухожилиям, и из-за их чрезвычайной чувствительности их можно активировать даже при дуновении воздуха на руку (Martin, 1985).
В дополнение к тельцам Пачини в голой коже имеется еще один быстро адаптирующийся рецептор. Большинство исследователей считают, что это тельца Мейснера, расположенные в дермальных сосочках кожи. Реагируя на низкочастотные колебания от 2 до 40 Гц, этот рецептор состоит из терминальных ветвей миелинизированного нервного волокна среднего размера, покрытых одним или несколькими слоями, по-видимому, модифицированных шванновских клеток, называемых ламинарными клетками. Нейриты и ламинарные клетки рецептора могут соединяться с базальной клеткой в эпидермисе (рис. 5).
Рисунок 5. Тельца Мейснера представляют собой слабо инкапсулированные сенсорные рецепторы в дермальных сосочках гладкой кожи.

Если тельца Мейснера выборочно инактивировать инъекцией местного анестетика через кожу, ощущение трепетания или низкочастотной вибрации теряется. Это говорит о том, что он функционально дополняет высокочастотную способность тельца Пачини. Вместе эти два рецептора обеспечивают нейронные сигналы, достаточные для того, чтобы объяснить человеческую чувствительность к полному диапазону вибраций (Mountcastle et al., 1967).
Кожные рецепторы, связанные со свободными нервными окончаниями
В дерме обнаруживается множество до сих пор не идентифицируемых миелиновых и немиелинизированных волокон. Большое количество только проходит на своем пути к коже, мышцам или надкостнице, в то время как другие (как миелинизированные, так и немиелинизированные), по-видимому, заканчиваются в дерме. За некоторыми исключениями, такими как тельца Пачини, большинство волокон в дерме заканчиваются плохо определенными способами или просто в виде свободных нервных окончаний.
Хотя для дифференциации этих плохо определенных окончаний необходимы дополнительные анатомические исследования, физиологические исследования ясно показали, что эти волокна кодируют множество событий окружающей среды. Например, свободные нервные окончания, обнаруженные на стыке между дермой и эпидермисом, отвечают за кодирование стимулов окружающей среды, которые интерпретируются как холод, тепло, тепло, боль, зуд и щекотание. Пока неизвестно, какие из этих разных классов тонких волокон передают те или иные ощущения.
Очевидное анатомическое сходство этих свободных нервных окончаний, вероятно, связано с ограничениями наших методов исследования, поскольку постепенно выявляются структурные различия между свободными нервными окончаниями. Например, в голой коже различают два различных терминальных типа свободных нервных окончаний: толстый, короткий рисунок и длинный, тонкий. Исследования волосистой кожи человека продемонстрировали наличие гистохимически распознаваемых нервных окончаний, которые заканчиваются на границе дермы и эпидермиса: пенициллезные и сосочковые окончания. Первые возникают из немиелинизированных волокон и образуют сеть окончаний; Напротив, последние возникают из миелинизированных волокон и заканчиваются вокруг устьев волос, как упоминалось ранее. Предположительно, эти структурные различия соответствуют функциональным различиям.
Хотя пока невозможно приписать определенные функции отдельным структурным единицам, из физиологических экспериментов ясно, что существуют функционально различные категории свободных нервных окончаний. Было обнаружено, что одно маленькое миелиновое волокно реагирует на холод у людей. Еще одно немиелинизированное волокно, обслуживающее свободные нервные окончания, реагирует на тепло. Каким образом один класс свободных нервных окончаний может избирательно реагировать на понижение температуры, а повышение температуры кожи может провоцировать другой класс на передачу сигнала о тепле, неизвестно. Исследования показывают, что активация одного небольшого волокна со свободным окончанием может быть ответственна за ощущения зуда или щекотки, в то время как считается, что существуют два класса маленьких волокон, особенно чувствительных к вредным механическим и вредным химическим или термическим раздражителям, обеспечивающим нервную основу для покалывания. жгучая боль (Keele, 1964).
Окончательная корреляция между анатомией и физиологической реакцией ожидает разработки более совершенных методов. Это один из основных камней преткновения в лечении таких расстройств, как каузалгия, парестезия и гиперпатия, которые продолжают представлять дилемму для врача.
Повреждение периферического нерва
Нейронную функцию можно разделить на две категории: сенсорную и моторную. Повреждение периферического нерва, обычно возникающее в результате сдавления или разрыва нерва, может привести к нарушению либо функции, либо того и другого, в зависимости от типа волокон в поврежденном нерве. Некоторые аспекты потери моторики, как правило, неправильно интерпретируются или игнорируются, поскольку эти сигналы не идут к мышцам, а скорее влияют на вегетативную сосудистую регуляцию, регуляцию температуры, характер и толщину эпидермиса и состояние кожных механорецепторов. Потеря двигательной иннервации здесь не будет обсуждаться, равно как и потеря иннервации, влияющая на другие органы чувств, кроме тех, которые отвечают за кожную чувствительность.
Потеря сенсорной иннервации кожи создает уязвимость для дальнейшего повреждения, поскольку оставляет анестезированную поверхность, которая не способна сигнализировать о потенциально вредных раздражителях. После повреждения анестезированные поверхности кожи заживают медленно, возможно, отчасти из-за отсутствия вегетативной иннервации, которая обычно регулирует такие ключевые факторы, как регуляция температуры и клеточное питание.
В течение нескольких недель денервированные кожные сенсорные рецепторы начинают атрофироваться, процесс, который легко наблюдать в крупных инкапсулированных рецепторах, таких как тельца Пачини и Мейснера. Если может произойти регенерация аксонов, за этим может последовать восстановление функции, но качество восстановленной функции будет зависеть от характера первоначального повреждения и продолжительности денервации (McKinnon and Dellon, 1988).
Восстановление после повреждения нерва происходит быстрее, намного полнее и функциональнее, чем восстановление после разрыва нерва. Два фактора объясняют благоприятный прогноз при защемлении нерва. Во-первых, большее количество аксонов может снова достичь контакта с кожей, чем после пересечения; во-вторых, соединения направляются обратно в исходное место благодаря шванновским клеткам и выстилкам, известным как базальные мембраны, обе из которых остаются интактными в раздавленном нерве, тогда как после пересечения нерва нервы часто перемещаются в неправильные участки поверхности кожи, следуя за неправильные пути шванновских клеток. Последняя ситуация приводит к тому, что искаженная пространственная информация отправляется в соматосенсорную кору головного мозга. Однако в обоих случаях регенерирующие аксоны, по-видимому, способны вернуться к тому же классу сенсорных рецепторов, которые они ранее обслуживали.
Реиннервация кожного рецептора — постепенный процесс. Когда растущий аксон достигает поверхности кожи, рецептивные поля становятся меньше, чем обычно, а пороговое значение выше. Эти рецептивные точки со временем расширяются и постепенно сливаются в более крупные поля. Чувствительность к механическим раздражителям становится выше и часто приближается к чувствительности нормальных сенсорных рецепторов этого класса. Исследования с использованием стимулов постоянного прикосновения, движущегося прикосновения и вибрации показали, что сенсорные модальности, связанные с различными типами рецепторов, возвращаются к анестезированным областям с разной скоростью.
Под микроскопом видно, что денервированная голая кожа тоньше, чем в норме, с уплощенными эпидермальными гребнями и меньшим количеством слоев клеток. Это подтверждает, что нервы имеют трофическое или питательное влияние на кожу. Вскоре после восстановления иннервации гребни дермы становятся лучше развиты, эпидермис утолщается, и можно обнаружить аксоны, проникающие через базальную мембрану. По мере того, как аксон возвращается к тельцу Мейснера, тельце начинает увеличиваться в размерах, и ранее уплощенная, атрофическая структура возвращается к исходной форме. Если денервация была длительной, новое тельце может образоваться рядом с исходным атрофическим скелетом, который остается денервированным (Dellon, 1981).
Как видно, понимание последствий повреждения периферических нервов требует знания нормального функционирования, а также степени функционального восстановления. В то время как эта информация доступна для некоторых нервных клеток, другие требуют дальнейшего изучения, оставляя ряд темных областей в нашем понимании роли кожных нервов в здоровье и болезни.