Figure 1. The female reproductive system.
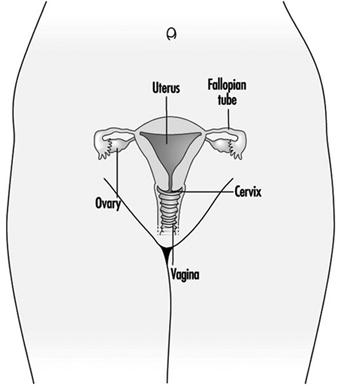
The female reproductive system is controlled by components of the central nervous system, including the hypothalamus and pituitary. It consists of the ovaries, the fallopian tubes, the uterus and the vagina (Figure 1). The ovaries, the female gonads, are the source of oocytes and also synthesize and secrete oestrogens and progestogens, the major female sex hormones. The fallopian tubes transport oocytes to and sperm from the uterus. The uterus is a pear-shaped muscular organ, the upper part of which communicates through the fallopian tubes to the abdominal cavity, while the lower part is contiguous through the narrow canal of the cervix with the vagina, which passes to the exterior. Table 1 summarizes compounds, clinical manifestations, site and mechanisms of action of potential reproductive toxicants.
Table 1. Potential female reproductive toxicants
| Compound | Clinical manifestation | Site | Mechanism/target |
| Chemical reactivity | |||
| Alkylating agents |
Altered menses Amenorrhoea Ovarian atrophy Decreased fertility Premature menopause |
Ovary Uterus |
Granulosa cell cytotoxicity Oocyte cytotoxicity Endometrial cell cytotoxicity |
| Lead | Abnormal menses Ovarian atrophy Decreased fertility |
Hypothalamus Pituitary Ovary |
Decreased FSH Decreased progesterone |
| Mercury | Abnormal menses | Hypothalamus Ovary |
Altered gonadotrophin production and secretion Follicle toxicity Granulosa cell proliferation |
| Cadmium | Follicular atresia Persistent diestrus |
Ovary Pituitary Hypothalamus |
Vascular toxicity Granulosa cell cytotoxicity Cytotoxicity |
| Structural similarity | |||
| Azathioprine | Reduced follicle numbers | Ovary Oogenesis |
Purine analog Disruption of DNA/RNA synthesis |
| Chlordecone | Impaired fertility | Hypothalamus | Oestrogen agonist |
| DDT | Altered menses | Pituitary | FSH, LH disruption |
| 2,4-D | Infertility | ||
| Lindane | Amenorrhoea | ||
| Toxaphene | Hypermenorrhoea | ||
| PCBs, PBBs | Abnormal menses | FSH, LH disruption | |
Source: From Plowchalk, Meadows and Mattison 1992. These compounds are suggested to be direct-acting reproductive toxicants based primarily on toxicity testing in experimental animals.
The Hypothalamus and Pituitary
The hypothalamus is located in the diencephalon, which sits on top of the brainstem and is surrounded by the cerebral hemispheres. The hypothalamus is the principal intermediary between the nervous and the endocrine systems, the two major control systems of the body. The hypothalamus regulates the pituitary gland and hormone production.
The mechanisms by which a chemical might disrupt the reproductive function of the hypothalamus generally include any event that could modify the pulsatile release of gonadotrophin releasing hormone (GnRH). This may involve an alteration in either the frequency or the amplitude of GnRH pulses. The processes susceptible to chemical injury are those involved in the synthesis and secretion of GnRH—more specifically, transcription or translation, packaging or axonal transport, and secretory mechanisms. These processes represent sites where direct-acting chemically reactive compounds might interfere with hypothalmic synthesis or release of GnRH. An altered frequency or amplitude of GnRH pulses could result from disruptions in stimulatory or inhibitory pathways that regulate the release of GnRH. Investigations of the regulation of the GnRH pulse generator have shown that catecholamines, dopamine, serotonin, γ-aminobutyric acid, and endorphins all have some potential for altering the release of GnRH. Therefore, xenobiotics that are agonists or antagonists of these compounds could modify GnRH release, thus interfering with communication with the pituitary.
Prolactin, follicle-stimulating hormone (FSH) and luteinizing hormone (LH) are three protein hormones secreted by the anterior pituitary that are essential for reproduction. These play a critical role in maintaining the ovarian cycle, governing follicle recruitment and maturation, steroidogenesis, completion of ova maturation, ovulation and luteinization.
The precise, finely tuned control of the reproductive system is accomplished by the anterior pituitary in response to positive and negative feedback signals from the gonads. The appropriate release of FSH and LH during the ovarian cycle controls normal follicular development, and the absence of these hormones is followed by amenorrhoea and gonadal atrophy. The gonadotrophins play a critical role in initiating changes in the morphology of ovarian follicles and in their steroidal microenvironments through the stimulation of steroid production and the induction of receptor populations. Timely and adequate release of these gonadotrophins is also essential for ovulatory events and a functional luteal phase. Because gonadotrophins are essential for ovarian function, altered synthesis, storage or secretion may seriously disrupt reproductive capacity. Interference with gene expression—whether in transcription or translation, post-translational events or packaging, or secretory mechanisms—may modify the level of gonadotrophins reaching the gonads. Chemicals that act by means of structural similarity or altered endocrine homeostasis might produce effects by interference with normal feedback mechanisms. Steroid-receptor agonists and antagonists might initiate an inappropriate release of gonadotrophins from the pituitary, thereby inducing steroid-metabolizing enzymes, reducing steroid half-life and subsequently the circulating level of steroids reaching the pituitary.
The Ovary
The ovary in primates is responsible for the control of reproduction through its principal products, oocytes and steroid and protein hormones. Folliculogenesis, which involves both intraovarian and extraovarian regulatory mechanisms, is the process by which oocytes and hormones are produced. The ovary itself has three functional subunits: the follicle, the oocyte and the corpus luteum. During the normal menstrual cycle, these components, under the influence of FSH and LH, function in concert to produce a viable ovum for fertilization and a suitable environment for implantation and subsequent gestation.
During the preovulatory period of the menstrual cycle, follicle recruitment and development occur under the influence of FSH and LH. The latter stimulates the production of androgens by thecal cells, whereas the former stimulates the aromatization of androgens into oestrogens by the granulosa cells and the production of inhibin, a protein hormone. Inhibin acts at the anterior pituitary to decrease the release of FSH. This prevents excess stimulation of follicular development and allows continuing development of the dominant follicle—the follicle destined to ovulate. Oestrogen production increases, stimulating both the LH surge (resulting in ovulation) and the cellular and secretory changes in the vagina, cervix, uterus and oviduct that enhance spermatozoa viability and transport.
In the postovulatory phase, thecal and granulosa cells remaining in the follicular cavity of the ovulated ovum, form the corpus luteum and secrete progesterone. This hormone stimulates the uterus to provide a proper environment for implantation of the embryo if fertilization occurs. Unlike the male gonad, the female gonad has a finite number of germ cells at birth and is therefore uniquely sensitive to reproductive toxicants. Such exposure of the female can lead to decreased fecundity, increased pregnancy wastage, early menopause or infertility.
As the basic reproductive unit of the ovary, the follicle maintains the delicate hormonal environment necessary to support the growth and maturation of an oocyte. As previously noted, this complex process is known as folliculogenesis and involves both intraovarian and extraovarian regulation. Numerous morphological and biochemical changes occur as a primordial follicle progresses to a pre-ovulatory follicle (which contains a developing oocyte), and each stage of follicular growth exhibits unique patterns of gonadotrophin sensitivity, steroid production and feedback pathways. These characteristics suggest that a number of sites are available for xenobiotic interaction. Also, there are different follicle populations within the ovary, which further complicates the situation by allowing for differential follicle toxicity. This creates a situation in which the patterns of infertility induced by a chemical agent would depend on the follicle type affected. For example, toxicity to primordial follicles would not produce immediate signs of infertility but would ultimately shorten the reproductive lifespan. On the other hand, toxicity to antral or preovulatory follicles would result in an immediate loss of reproductive function. The follicle complex is composed of three basic components: granulosa cells, thecal cells and the oocyte. Each of these components has characteristics that may make it uniquely susceptible to chemical injury.
Several investigators have explored methodology for screening xenobiotics for granulosa cell toxicity by measuring the effects on progesterone production by granulosa cells in culture. Oestradiol suppression of progesterone production by granulosa cells has been utilized to verify granulosa cell responsiveness. The pesticide p,p’-DDT and its o,p’-DDT isomer produce supression of progesterone production apparently with potencies equal to that of oestradiol. By contrast, the pesticides malathion, arathion and dieldrin and the fungicide hexachlorobenzene are without effect. Further detailed analysis of isolated granulosa cell responses to xenobiotics is needed to define the utility of this assay system. The attractiveness of isolated systems such as this is economy and ease of use; however, it is important to remember that granulosa cells represent only one component of the reproductive system.
Thecal cells provide precursors for steroids synthesized by granulosa cells. Thecal cells are believed to be recruited from ovarian stroma cells during follicle formation and growth. Recruitment may involve stromal cellular proliferation as well as migration to regions around the follicle. Xenobiotics that impair cell proliferation, migration and communication will impact on thecal cell function. Xenobiotics that alter thecal androgen production may also impair follicle function. For example, the androgens metabolized to oestrogens by granulosa cells are provided by thecal cells. Alterations in thecal cell androgen production, either increases or decreases, are expected to have a significant effect on follicle function. For example, it is believed that excess production of androgens by thecal cells will lead to follicle atresia. In addition, impaired production of androgens by thecal cells may lead to decreased poestrogen production by granulosa cells. Either circumstance will clearly impact on reproductive performance. At resent, little is known about thecal cell vulnerability to xenobiotics.
Although there is a acuity of information defining the vulnerability of ovarian cells to xenobiotics, there are data clearly demonstrating that oocytes can be damaged or destroyed by such agents. Alkylating agents destroy oocytes in humans and experimental animals. Lead produces ovarian toxicity. Mercury and cadmium also produce ovarian damage that may be mediated through oocyte toxicity.
Fertilization to Implantation
Gametogenesis, release and union of male and female germ cells are all preliminary events leading to a zygote. Sperm cells deposited in the vagina must enter the cervix and move through the uterus and into the fallopian tube to meet the ovum. penetration of ovum by sperm and the merging of their respective DNA comprise the process of fertilization. After fertilization cell division is initiated and continues during the next three or four days, forming a solid mass of cells called a morula. The cells of the morula continue to divide, and by the time the developing embryo reaches the uterus it is a hollow ball called a blastocyst.
Following fertilization, the developing embryo migrates through the fallopian tube into the uterus. The blastocyst enters the uterus and implants in the endometrium approximately seven days after ovulation. At this time the endometrium is in the postovulatory phase. Implantation enables the blastocyst to absorb nutrients or toxicants from the glands and blood vessels of the endometrium.