皮膚敏感性共享所有基本感官的主要元素。 外部世界的特性,如顏色、聲音或振動,被稱為感覺受體的特殊神經細胞末梢接收,將外部數據轉化為神經衝動。 然後這些信號被傳送到中樞神經系統,在那裡它們成為解釋我們周圍世界的基礎。
認識到這些過程的三個要點是很有用的。 首先,能量和能量水平的變化只能由能夠檢測所討論的特定能量類型的感覺器官感知。 (這就是為什麼微波、X 射線和紫外線都是危險的;我們沒有能力檢測它們,因此即使在致命的水平上它們也不會被感知。)其次,我們的感知必然是現實的不完美影子,因為我們的中心神經系統僅限於根據其感覺受體傳遞的信號重建不完整的圖像。 第三,我們的感覺系統為我們提供了比靜態條件更準確的關於環境變化的信息。 例如,我們配備了對閃爍的燈光敏感的感覺感受器,或者對微風引起的微小溫度波動敏感; 我們沒有足夠的能力接收有關穩定溫度或皮膚上恆定壓力的信息。
傳統上,皮膚感覺分為兩類:皮膚和深層。 雖然深層敏感性依賴於位於肌肉、肌腱、關節和骨膜(骨骼周圍的膜)中的受體,但我們在此關注的皮膚敏感性處理皮膚受體接收的信息:具體來說,各種類別的位於真皮和表皮交界處或附近的皮膚感受器。
將皮膚感受器連接到中樞神經系統的所有感覺神經都具有大致相同的結構。 細胞的大體存在於一組其他神經細胞體中,稱為神經節,位於脊髓附近,並通過細胞幹的一個狹窄分支連接到脊髓,稱為軸突。 大多數起源於脊髓的神經細胞或神經元將軸突發送到骨骼、肌肉、關節,或者在皮膚敏感的情況下發送到皮膚。 就像絕緣電線一樣,每個軸突沿其路線和末端都覆蓋著稱為雪旺氏細胞的細胞保護層。 這些雪旺細胞產生一種稱為髓磷脂的物質,它像鞘一樣包裹著軸突。 沿途的間隔是髓鞘中的微小斷裂,稱為郎飛結。 最後,在軸突的末端發現了專門接收和重新傳輸有關外部環境的信息的組件:感覺受體 (Mountcastle 1974)。
不同類別的皮膚感受器,就像所有的感覺感受器一樣,以兩種方式定義:通過它們的解剖結構,以及它們沿著神經纖維發送的電信號的類型。 結構獨特的受體通常以其發現者的名字命名。 皮膚中發現的感覺受體種類相對較少,可分為三大類:機械感受器、熱感受器和傷害感受器。
所有這些受體只有在首先用一種電化學神經語言對特定刺激進行編碼後,才能傳達有關特定刺激的信息。 這些神經代碼使用不同頻率和模式的神經衝動,科學家們才剛剛開始破譯這些神經衝動。 事實上,神經生理學研究的一個重要分支完全致力於研究感覺受體以及它們將環境中的能量狀態轉化為神經代碼的方式。 一旦生成代碼,它們就會沿著傳入纖維集中傳送,傳入纖維是通過將信號傳送到中樞神經系統來為受體服務的神經細胞。
受體產生的信息可以根據對連續的、不變的刺激的反應進行細分:緩慢適應的受體在恆定刺激的持續時間內向中樞神經系統發送電化學脈衝,而快速適應的受體逐漸減少它們的放電穩定刺激的存在,直到它們達到低基線水平或完全停止,然後停止通知中樞神經系統有關刺激的持續存在。
因此,疼痛、溫暖、寒冷、壓力和振動等截然不同的感覺是由不同類別的感覺受體及其相關神經纖維的活動產生的。 例如,術語“顫動”和“振動”用於區分由兩類不同類型的振動敏感受體編碼的兩種略有不同的振動感覺 (Mountcastle et al. 1967)。 被稱為刺痛、灼痛和酸痛的三種重要的痛覺類別都與一類不同的傷害性傳入纖維有關。 然而,這並不是說特定的感覺必然只涉及一類感受器。 不止一種受體類別可能對給定的感覺有貢獻,事實上,感覺可能會因不同受體類別的相對貢獻而有所不同 (Sinclair 1981)。
上述總結基於皮膚感覺功能的特異性假說,該假說首先由一位名叫 Von Frey 的德國醫生於 1906 年提出。儘管在過去的一個世紀中至少提出了兩個同等或可能更受歡迎的其他理論,但 Von Frey 的假說已經現在得到了事實證據的有力支持。
對持續的皮膚壓力有反應的受體
在手部,相對較大的有髓纖維(直徑 5 至 15 毫米)從稱為乳頭下神經叢的皮下神經網絡發出,終止於真皮和表皮交界處的一束神經末梢(圖 1)。 在毛茸茸的皮膚中,這些神經末梢形成可見的表面結構,稱為 觸摸屏; 在無毛或無毛的皮膚中,神經末梢位於皮脊的底部(例如形成指紋的皮脊)。 在那裡,在觸摸圓頂中,每個神經纖維尖端或神經突都被一種特殊的上皮細胞包圍,稱為 默克爾細胞 (見圖 2 和 3)。
圖 1. 皮膚橫截面示意圖
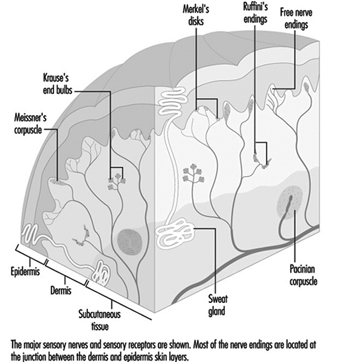
圖 2. 每個凸起皮膚區域的觸摸圓頂包含 30 到 70 個默克爾細胞。
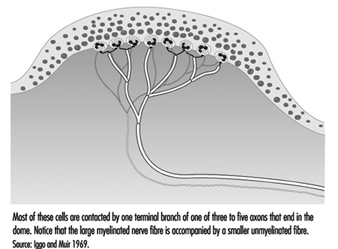
圖 3. 在電子顯微鏡的更高放大倍數下,可以看到默克爾細胞(一種特殊的上皮細胞)附著在將表皮與真皮分開的基底膜上。
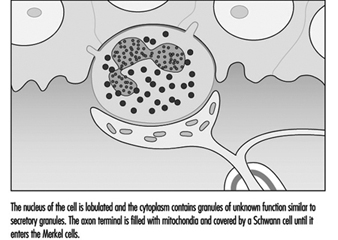
Merkel 細胞神經突複合體將機械能轉化為神經衝動。 雖然對細胞的作用或其轉導機制知之甚少,但它已被確定為一種緩慢適應的受體。 這意味著在包含 Merkel 細胞的觸摸圓頂上施加壓力會導致受體在刺激期間產生神經衝動。 這些脈衝的頻率與刺激強度成正比,從而告知大腦皮膚壓力的持續時間和大小。
與默克爾細胞一樣,第二個緩慢適應的受體也通過發出穩定皮膚壓力的大小和持續時間信號來為皮膚服務。 只有通過顯微鏡才能看到,這個受體,被稱為 魯菲尼受體,由一組從有髓纖維中出現並被結締組織細胞包裹的神經突組成。 膠囊結構內的纖維明顯將局部皮膚變形傳遞給神經突,神經突又產生沿著神經高速公路發送到中樞神經系統的信息。 對皮膚的壓力導致神經衝動的持續釋放; 與默克爾細胞一樣,神經衝動的頻率與刺激強度成正比。
儘管它們有相似之處,但默克爾細胞和魯菲尼受體之間存在一個顯著差異。 當魯菲尼受體受到刺激時會產生感覺,而容納默克爾細胞的觸摸圓頂的刺激不會產生有意識的感覺; 因此,觸摸圓頂是一個神秘的受體,因為它在神經功能中的實際作用仍然未知。 因此,Ruffini 受體被認為是唯一能夠提供壓力感官體驗或持續觸摸所必需的神經信號的受體。 此外,已經表明,緩慢適應的 Ruffini 受體解釋了人類根據強度等級對皮膚壓力進行評級的能力。
對振動和皮膚運動有反應的受體
與緩慢適應的機械感受器相反,快速適應的感受器在持續的皮膚壓痕過程中保持沉默。 然而,它們非常適合發出振動和皮膚運動信號。 注意到兩個一般類別: 那些在毛茸茸的皮膚上,與個別毛髮相關; 以及那些在無毛或無毛的皮膚上形成微粒末端的。
毛髮受體
一根典型的頭髮被神經末梢網絡包裹著,神經末梢從五到九個大的有髓軸突分支出來(圖 4)。 在靈長類動物中,這些末端分為三類:披針形末端、紡錘狀末端和乳頭狀末端。 這三者都在迅速適應,因此只有在運動發生時,頭髮的穩定偏轉才會引起神經衝動。 因此,這些感受器對移動或振動刺激非常敏感,但很少或根本不提供有關壓力或持續觸摸的信息。
圖 4. 毛幹是檢測運動的神經末梢平台。
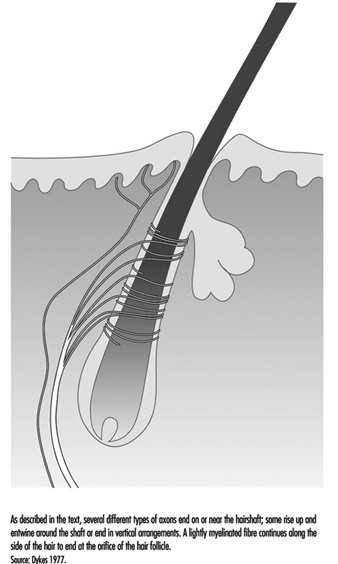
披針形的末端源自在毛髮周圍形成網狀結構的重度髓鞘纖維。 末端神經突失去了它們通常對雪旺細胞的覆蓋,並在毛髮底部的細胞中發揮作用。
紡錘狀末端由被雪旺細胞包圍的軸突末端形成。 末端上升到傾斜的毛幹,並在皮脂腺或產油腺下方的半圓形簇中結束。 乳頭狀末梢與紡錘狀末端不同,因為它們不是終止於毛幹,而是終止於毛髮孔周圍的游離神經末梢。
據推測,在毛髮上發現的受體類型之間存在功能差異。 這可以部分地從神經末端在毛幹上的方式的結構差異和部分從軸突直徑的差異推斷出來,因為不同直徑的軸突連接到不同的中央中繼區域。 儘管如此,多毛皮膚中受體的功能仍然是一個有待研究的領域。
無毛皮膚中的受體
受體的解剖結構與其產生的神經信號的相關性在具有小體或封裝末端的大型且易於操縱的受體中最為明顯。 特別容易理解的是 pacininan 和 Meissner 小體,它們就像上面討論的毛髮中的神經末梢一樣,傳遞振動的感覺。
pacinian 小體足夠大,可以用肉眼看到,因此很容易將受體與特定的神經反應聯繫起來。 它位於真皮中,通常圍繞肌腱或關節,呈洋蔥狀結構,大小為 0.5 × 1.0 毫米。 它由身體最大的傳入纖維之一提供服務,直徑為 8 至 13 微米,傳導速度為每秒 50 至 80 米。 它的解剖結構,通過光學和電子顯微鏡進行了充分研究,是眾所周知的。
小體的主要成分是由包圍充滿液體的空間的細胞材料形成的外核。 然後外核本身被一個被中央管和毛細管網絡穿透的膠囊包圍。 穿過管的是一根直徑為 7 至 11 毫米的單根有髓神經纖維,它變成一個長的無髓神經末梢,深入到小體的中心。 末端軸突是橢圓形的,具有分支狀突起。
pacinian 小體是一種快速適應受體。 因此,當受到持續壓力時,它只會在刺激的開始和結束時產生衝動。 它對高頻振動(80 至 400 赫茲)有反應,對 250 赫茲左右的振動最敏感。 通常,這些感受器會對沿骨骼和肌腱傳輸的振動做出反應,並且由於它們極其敏感,它們可能被手上的一團空氣激活 (Martin 1985)。
除了 pacinian 小體外,無毛皮膚中還有另一種快速適應受體。 大多數研究人員認為它是位於皮膚真皮乳頭中的邁斯納小體。 該受體對 2 至 40 赫茲的低頻振動有反應,由中等大小的有髓神經纖維的末端分支組成,該神經纖維被包裹在一層或幾層似乎經過修飾的雪旺細胞(稱為層狀細胞)中。 受體的神經突和層狀細胞可能與表皮中的基底細胞相連(圖 5)。
圖 5. 邁斯納小體是無毛皮膚真皮乳頭中鬆散包裹的感覺受體。
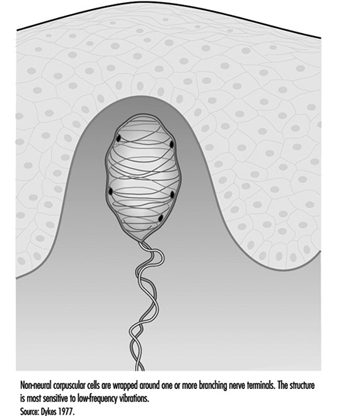
如果通過皮膚注射局部麻醉劑選擇性地使邁斯納小體失活,那麼顫動或低頻振動的感覺就會消失。 這表明它在功能上補充了 pacinian 小體的高頻能力。 這兩個受體一起提供的神經信號足以解釋人類對各種振動的敏感性 (Mountcastle et al. 1967)。
與游離神經末梢相關的皮膚受體
在真皮中發現了許多仍然無法辨認的有髓和無髓纖維。 大量只是通過,在前往皮膚、肌肉或骨膜的途中,而其他(有髓鞘和無髓鞘)似乎終止於真皮。 除了少數例外,例如 pacinian 小體,真皮中的大多數纖維似乎以不明確的方式結束或簡單地作為游離神經末梢。
雖然需要更多的解剖學研究來區分這些不明確的末端,但生理學研究已經清楚地表明,這些纖維編碼了各種環境事件。 例如,在真皮和表皮交界處發現的游離神經末梢負責編碼環境刺激,這些刺激將被解釋為冷、暖、熱、痛、癢和癢。 目前尚不清楚這些不同類別的小纖維中的哪一種能傳達特定的感覺。
這些游離神經末梢在解剖學上的明顯相似性可能是由於我們調查技術的局限性,因為游離神經末梢之間的結構差異正在慢慢被發現。 例如,在無毛皮膚中,已區分出兩種不同的游離神經末梢末端模式:厚而短的模式和長而薄的模式。 對人類多毛皮膚的研究表明,組織化學可識別的神經末梢終止於真皮-表皮交界處:青黴狀末梢和乳頭狀末梢。 前者產生於無髓纖維並形成末端網絡; 相反,如前所述,後者起源於有髓纖維並在毛孔周圍結束。 據推測,這些結構差異對應於功能差異。
雖然還不可能將特定功能分配給單個結構實體,但從生理學實驗可以清楚地看出,存在功能不同的游離神經末梢類別。 已發現一種小的有髓纖維對人類的寒冷有反應。 另一種服務於游離神經末梢的無髓纖維對溫暖有反應。 一類游離神經末梢如何選擇性地對溫度下降做出反應,而皮膚溫度的升高如何激發另一類神經末梢來發出溫暖信號尚不清楚。 研究表明,激活一根具有自由末端的小纖維可能會導致瘙癢或發癢的感覺,而據信有兩類小纖維對有害機械和有害化學或熱刺激特別敏感,為刺痛提供了神經基礎和灼痛 (Keele 1964)。
解剖學和生理反應之間的明確相關性等待更先進技術的發展。 這是治療灼痛、感覺異常和痛覺過敏等疾病的主要障礙之一,這些疾病一直困擾著醫生。
周圍神經損傷
神經功能可分為兩類:感覺和運動。 周圍神經損傷通常由神經擠壓或切斷引起,可損害其中一種或兩種功能,具體取決於受損神經中的纖維類型。 運動喪失的某些方面往往會被誤解或忽視,因為這些信號不會傳遞到肌肉,而是會影響自主血管控制、溫度調節、表皮的性質和厚度以及皮膚機械受體的狀況。 這裡不會討論運動神經支配的喪失,神經支配的喪失也不會影響負責皮膚感覺的感官。
皮膚失去感覺神經支配會造成進一步傷害的脆弱性,因為它會留下無法發出潛在有害刺激信號的麻醉表面。 一旦受傷,麻醉的皮膚表面癒合緩慢,部分原因可能是缺乏通常調節溫度調節和細胞營養等關鍵因素的自主神經支配。
在幾週的時間裡,去神經支配的皮膚感覺受體開始萎縮,這一過程很容易在大的封裝受體(如 pacinian 和 Meissner 小體)中觀察到。 如果軸突可以再生,功能可能會隨之恢復,但恢復功能的質量將取決於原始損傷的性質和去神經支配的持續時間 (McKinnon 和 Dellon 1988)。
與神經切斷後的恢復相比,神經損傷後的恢復更快、更完整、功能更強大。 有兩個因素可以解釋神經損傷的良好預後。 首先,與橫斷後相比,更多的軸突可能再次與皮膚接觸; 其次,這些連接被雪旺細胞和被稱為基底膜的襯裡引導回到它們的原始位置,這兩者在被壓碎的神經中都保持完整,而在神經橫斷後,神經通常通過跟隨錯誤的雪旺細胞路徑。 後一種情況導致扭曲的空間信息被發送到大腦的體感皮層。 然而,在這兩種情況下,再生的軸突似乎都能夠找到返回它們之前服務的同一類感覺受體的途徑。
皮膚受體的神經再支配是一個漸進的過程。 隨著生長的軸突到達皮膚表面,感受野比正常情況下更小,而閾值更高。 這些接受點隨時間擴展並逐漸合併成更大的場。 對機械刺激的敏感性變得更高,並且通常接近該類別的正常感覺受體的敏感性。 使用持續觸摸、移動觸摸和振動刺激的研究表明,歸因於不同類型受體的感覺方式以不同的速率返回麻醉區域。
在顯微鏡下觀察,去神經支配的無毛皮膚比正常皮膚更薄,具有扁平的表皮脊和更少的細胞層。 這證實了神經對皮膚有營養或營養影響。 神經支配恢復後不久,真皮脊變得更好,表皮變得更厚,並且可以發現軸突穿透基底膜。 隨著軸突回到邁斯納小體,小體開始變大,之前扁平、萎縮的結構恢復到原來的形式。 如果去神經支配持續時間很長,一個新的小體可能會在原始萎縮的骨骼附近形成,而該骨骼仍處於去神經支配狀態(Dellon 1981)。
可以看出,了解周圍神經損傷的後果需要了解正常功能以及功能恢復的程度。 雖然此信息可用於某些神經細胞,但其他神經細胞需要進一步研究,從而在我們對皮神經在健康和疾病中的作用的理解中留下許多模糊區域。