人類一生都生活在一個非常小的、受到嚴密保護的體內溫度範圍內。 活細胞的最大耐受範圍從大約 0ºC(冰晶形成)到大約 45ºC(細胞內蛋白質的熱凝固); 然而,人類只能在非常短的時間內忍受低於 35ºC 或高於 41ºC 的內部溫度。 為了將內部溫度保持在這些限制範圍內,人們已經對急性熱應激產生了非常有效且在某些情況下專門的生理反應。 這些反應——旨在促進身體熱量的保存、產生或消除——涉及多個身體系統的精細控制協調。
人體熱平衡
到目前為止,傳遞給身體的最大熱源來自代謝熱的產生 (M). 即使在最高機械效率下,肌肉工作所涉及的能量中有 75% 到 80% 會以熱量的形式釋放出來。 休息時,代謝率為 300 毫升 O2 每分鐘產生大約 100 瓦的熱負荷。 在耗氧量為 1 升/分鐘的穩態工作期間,會產生大約 350 W 的熱量——減去任何與外部工作相關的能量 (W). 即使在如此輕度至中度的工作強度下,如果沒有有效的散熱方式,身體核心溫度也會每 15 分鐘升高約 1,200 攝氏度。 事實上,非常健康的人可以產生超過 1 W 的熱量並持續 3 到 1984 小時而不會造成熱損傷 (Gisolfi and Wenger XNUMX)。
也可以通過輻射從環境中獲取熱量 (R) 和對流 (C) 如果地球溫度(輻射熱的量度)和空氣(幹球)溫度分別超過皮膚溫度。 這些熱增益途徑通常相對於 M,並且當皮膚與空氣的熱梯度反轉時,實際上成為熱損失的途徑。 熱量散失的最後途徑——蒸發 (E)—通常也是最重要的,因為汗液蒸發的潛熱很高——大約 680 Wh/l 蒸發的汗液。 這些關係將在本章的其他地方討論。
在涼爽到熱中性條件下,熱量增加與熱量損失平衡,沒有熱量儲存,體溫平衡; 那是:
M-W ± R ± C-E = 0
然而,在更嚴重的高溫下:
分子量–寬度±R±C >E
並儲存熱量。 特別是繁重的工作(高能量消耗會增加 M-W), 過高的空氣溫度 (增加 遙控+遙控), 高濕度 (這限制了 E) 和穿厚衣服或相對不透水的衣服(這會阻礙汗液的有效蒸發)會造成這種情況。 最後,如果運動時間延長或水分不足, E 可能會因身體排汗能力有限(短時間 1 至 2 升/小時)而超過。
體溫及其控制
為了描述對熱和冷的生理反應,身體分為兩個部分——“核心”和“外殼”。 核心溫度 (Tc) 表示內部或深部體溫,可以通過口腔、直腸或在實驗室環境中在食道或鼓膜(鼓膜)上測量。 外殼的溫度由平均皮膚溫度表示(Tsk). 人體的平均溫度 (Tb) 在任何時候都是這些溫度之間的加權平衡,即
Tb = k Tc + (1– k) Tsk
其中加權因子 k 從大約 0.67 到 0.90 不等。
當面臨熱中性挑戰(熱或冷應激)時,身體會努力控制 Tc 通過生理調整,和 Tc 向大腦提供主要反饋以協調這種控制。 雖然局部和平均皮膚溫度對於提供感官輸入很重要, Tsk 溫度隨環境溫度變化很大,在熱中性條件下平均約為 33 ºC,在炎熱的繁重工作條件下達到 36 至 37 ºC。 在全身和局部暴露於寒冷時,它會顯著下降; 觸覺靈敏度發生在 15 到 20 ºC 之間,而手靈巧性的臨界溫度在 12 到 16 ºC 之間。 上下痛閾值 Tsk 分別約為 43 ºC 和 10 ºC。
精確測繪研究已將最大體溫調節控制位置定位在稱為視前/前下丘腦 (POAH) 的大腦區域。 在這個區域是對加熱(熱敏感神經元)和冷卻(冷敏感神經元)都有反應的神經細胞。 該區域通過自主神經系統接收有關體溫的傳入感覺信息並向皮膚、肌肉和其他參與溫度調節的器官發送傳出信號,從而控制體溫。 中樞神經系統的其他區域(下丘腦後部、網狀結構、腦橋、延髓和脊髓)與 POAH 形成上行和下行連接,並發揮多種輔助功能。
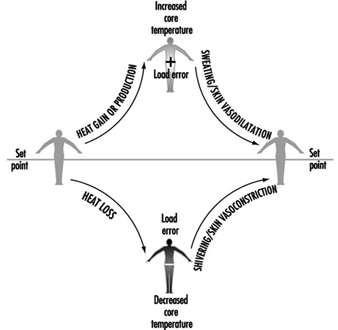
身體的控制系統類似於具有加熱和冷卻功能的房屋中的溫度恆溫控制。 當體溫升高到某個理論“設定點”溫度以上時,與冷卻(出汗、增加皮膚血流量)相關的效應器反應就會啟動。 當體溫低於設定點時,就會啟動熱增益反應(皮膚血流量減少、發抖)。 然而,與家庭供暖/製冷系統不同的是,人體體溫調節控制系統並不是簡單的開關係統,而是具有比例控制和變化率控制特性。 應當理解,“設定點溫度”僅存在於理論上,因此有助於形象化這些概念。 要全面了解與體溫調節設定點相關的機制,還有很多工作要做。
無論其基礎如何,設定點都是相對穩定的,並且不受工作或環境溫度的影響。 事實上,已知唯一改變設定點的劇烈擾動是參與發熱反應的一組內源性熱原。 身體為維持熱平衡而採用的效應器響應是響應“負載錯誤”而啟動和控制的,即體溫瞬時高於或低於設定點(圖 1)。 低於設定點的核心溫度會產生負負載誤差,導致熱量增加(皮膚顫抖、血管收縮)開始。 高於設定點的核心溫度會產生正負載誤差,導致熱損失效應器(皮膚血管擴張、出汗)被打開。 在每種情況下,由此產生的熱傳遞都會減少負載誤差,並有助於將體溫恢復到穩定狀態。
圖 1. 人體體溫調節模型。
高溫下的溫度調節
如上所述,人類主要通過乾燥(輻射和對流)和蒸發方式的組合向環境散失熱量。 為了促進這種交換,兩個主要的效應系統被打開和調節——皮膚血管擴張和出汗。 雖然皮膚血管擴張通常會導致乾燥(輻射和對流)熱損失的小幅增加,但它的主要功能是將熱量從核心傳遞到皮膚(內部熱傳遞),而汗液的蒸發提供了一種非常有效的冷卻血液的方法使其返回深層身體組織(外部熱傳遞)。
皮膚血管擴張
從核心傳遞到皮膚的熱量是皮膚血流量 (SkBF)、核心和皮膚之間的溫度梯度以及血液比熱(每升血液略低於 4 kJ/°C)的函數血)。 在熱中性環境中休息時,皮膚的血流量約為 200 至 500 毫升/分鐘,僅佔心臟泵出的總血液(心輸出量)的 5% 至 10%。 由於 4ºC 之間的梯度 Tc (約 37ºC)和 Tsk (此條件下約33ºC),人體為維持生命而產生的代謝熱不斷地對流至皮膚散發。 相比之下,在高溫條件下的高強度工作等嚴重高熱條件下,核心到皮膚的熱梯度較小,所需的熱傳遞是通過 SkBF 的大幅增加來完成的。 在最大熱應激下,SkBF 可達到 7 至 8 l/min,約為心輸出量的三分之一 (Rowell 1983)。 這種高血流量是通過一種鮮為人知的人類獨有機制實現的,這種機制被稱為“主動血管擴張系統”。 主動血管舒張涉及從下丘腦到皮膚小動脈的交感神經信號,但神經遞質尚未確定。
如上所述,SkBF 主要響應於 Tc 並且,在較小程度上, Tsk. Tc 隨著肌肉工作的開始和代謝熱的產生開始,上升,一旦達到某個閾值 Tc 達到時,SkBF 也開始急劇增加。 這種基本的體溫調節關係也受到非熱因素的影響。 第二級控制至關重要,因為當整體心血管穩定性受到威脅時,它會修改 SkBF。 皮膚中的靜脈非常順應,並且很大一部分循環體積匯集在這些血管中。 這通過減慢毛細管循環以增加傳輸時間來幫助熱交換; 然而,這種匯集,再加上出汗導致的體液流失,也可能會降低血液回流到心臟的速度。 在工作期間影響 SkBF 的非熱因素包括直立姿勢、脫水和正壓呼吸(呼吸器使用)。 這些通過反射起作用,當心臟充盈壓降低並且位於大靜脈和右心房中的牽張感受器被卸載時,這些反射被打開,因此在長時間的有氧運動中以直立姿勢最為明顯。 這些反射的作用是維持動脈壓,並在工作時維持足夠的血液流向活躍的肌肉。 因此,任何給定時間點的 SkBF 水平代表體溫調節和非體溫調節反射反應的綜合效應。
需要增加流向皮膚的血液以幫助調節溫度,這極大地影響了心血管系統調節血壓的能力。 因此,整個心血管系統對熱應激的協調反應是必要的。 會發生哪些心血管調整以增加皮膚流量和體積? 在涼爽或熱中性條件下工作期間,增加心率 (HR) 可以很好地支持所需的心輸出量增加,因為每搏輸出量 (SV) 的進一步增加是最小的,超過最大運動強度的 40%。 在高溫下,HR 在任何給定的工作強度下都較高,作為對降低的中心血容量 (CBV) 和 SV 的補償。 在更高水平的工作中,達到最大心率,因此這種心動過速無法維持必要的心輸出量。 身體提供高 SkBF 的第二種方式是將血流從肝臟、腎臟和腸道等區域分配出去 (Rowell 1983)。 這种血流重定向可以為皮膚提供額外的 800 至 1,000 毫升血流,並有助於抵消外周血池的不利影響。
抑汗療程
人體體溫調節汗液由 2 至 4 百萬個不均勻分佈在體表的小汗腺分泌。 與傾向於聚集(在面部和手部以及中軸和生殖器區域)並將汗液分泌到毛囊中的大汗腺不同,小汗腺將汗液直接分泌到皮膚表面。 這種汗液無味、無色且相對較稀,因為它是血漿的超濾液。 因此,它具有高蒸發潛熱,非常適合其冷卻目的。
作為這種冷卻系統有效性的一個例子,一個人在 2.3 升/分鐘的氧氣消耗下工作會產生淨代謝熱(M-W) 約 640 W。在沒有出汗的情況下,體溫每 1 至 6 分鐘會以約 7°C 的速度升高。 每分鐘約16克汗液的高效蒸發(合理速率),熱量散失速率與產熱速率相匹配,身體核心溫度可保持在穩定狀態; 那是,
M–W±R±C–E = 0
小汗腺結構簡單,由盤繞的分泌部分、導管和皮膚毛孔組成。 每個腺體產生的汗液量取決於腺體的結構和功能,而總出汗率又取決於腺體的募集(活性汗腺密度)和汗腺輸出量。 有些人比其他人出汗更多的事實主要歸因於汗腺大小的差異(Sato 和 Sato 1983)。 熱適應是汗液產生的另一個主要決定因素。 隨著年齡的增長,出汗率降低並不是因為激活的外分泌腺減少,而是因為每個腺體的排汗量減少(Kenney 和 Fowler 1988)。 這種下降可能與伴隨衰老過程的結構和功能改變有關。
與血管運動信號一樣,對汗腺的神經衝動起源於 POAH,並通過腦幹下降。 支配腺體的纖維是交感神經膽鹼能纖維,這是人體中罕見的組合。 雖然乙酰膽鹼是主要的神經遞質,但腎上腺素能遞質(兒茶酚胺)也會刺激外分泌腺。
在許多方面,控制出汗類似於控制皮膚血流。 兩者俱有相似的起病特徵(閾值)和與增加的線性關係 Tc. 背部和胸部往往出汗較早,局部出汗率與 Tc 對於這些站點來說是最陡峭的。 與 SkBF 一樣,出汗會受到非熱因素的影響,例如水合不足和高滲透壓。 同樣值得注意的是一種稱為“多汗症”的現象,它發生在非常潮濕的環境中或經常被濕衣服覆蓋的皮膚區域。 這些皮膚區域,由於它們持續處於潮濕狀態,會減少出汗量。 這是一種防止持續脫水的保護機制,因為留在皮膚上而不是蒸發的汗水沒有冷卻功能。
如果出汗率足夠,蒸發冷卻最終取決於濕潤皮膚和周圍空氣之間的水蒸氣壓力梯度。 因此,高濕度和厚重或不透水的衣服會限制蒸發冷卻,而乾燥的空氣、身體周圍的空氣流動和極少的多孔衣服會促進蒸發。 另一方面,如果工作繁重且大量出汗,蒸發冷卻同樣會受到身體排汗能力的限制(最大約 1 至 2 升/小時)。
寒冷時的溫度調節
與熱相比,人類對冷的反應方式的一個重要區別是,行為在對冷的體溫調節反應中起著更大的作用。 例如,在寒冷的環境條件下,穿著合適的衣服並採取能夠最大程度地減少可用於熱損失的表面積(“擠在一起”)的姿勢比在炎熱的環境條件下重要得多。 第二個區別是激素在冷應激期間發揮的作用更大,包括增加兒茶酚胺(去甲腎上腺素和腎上腺素)和甲狀腺激素的分泌。
皮膚血管收縮
防止身體通過輻射和對流散失熱量的有效策略是增加外殼提供的有效隔熱效果。 在人類中,這是通過減少流向皮膚的血流量——即通過皮膚血管收縮來實現的。 四肢的皮膚血管收縮比軀幹更明顯。 與主動血管舒張一樣,皮膚血管收縮也受交感神經系統控制,並受 Tc,Tsk 和當地的溫度。
皮膚降溫對心率和血壓反應的影響因被降溫的身體區域以及寒冷是否嚴重到足以引起疼痛而異。 例如,當雙手浸入冷水中時,心率、收縮壓(SBP)和舒張壓(DBP)都會升高。 當面部變冷時,由於廣泛的交感神經反應,SBP 和 DBP 會增加; 然而,由於副交感神經反射,HR 會下降 (LeBlanc 1975)。 為了進一步混淆對寒冷的整體反應的複雜性,一個人對另一個人的反應存在廣泛的差異。 如果冷應激足以降低身體核心溫度,HR 可能會增加(由於交感神經激活)或減少(由於中心血容量增加)。
感興趣的特定情況被稱為 寒冷引起的血管舒張 (CIVD)。 當手放在冷水中時,SkBF 最初會降低以保存熱量。 隨著組織溫度下降,SkBF 矛盾地增加,再次減少,並重複這種循環模式。 有人提出 CIVD 有利於防止組織因冷凍而受損,但這尚未得到證實。 從機制上講,當寒冷的直接影響嚴重到足以減少神經傳遞時,可能會發生短暫的擴張,這會暫時超越寒冷對血管交感神經受體的影響(介導收縮效應)。
戰栗
隨著身體冷卻的進行,第二道防線開始顫抖。 顫抖是表層肌肉纖維的隨機不自主收縮,它不會限制熱量損失,反而會增加熱量產生。 由於這種收縮不產生任何功,因此會產生熱量。 一個休息的人在劇烈顫抖時可以增加他或她的代謝熱產生大約三到四倍,並且可以增加 Tc 降低 0.5ºC。 引發顫抖的信號主要來自皮膚,除了大腦的 POAH 區域外,下丘腦後部也在很大程度上參與其中。
儘管許多個體因素會導致發抖(以及一般的耐寒性),但一個重要因素是身體肥胖。 皮下脂肪很少(2 至 3 毫米厚)的人在 40ºC 15 分鐘和 20ºC 10 分鐘後開始發抖,而絕緣脂肪較多(11 毫米)的人在 15ºC 和 60 分鐘後可能根本不會發抖在 10ºC (LeBlanc 1975)。