La tossicologia genetica, per definizione, è lo studio di come gli agenti chimici o fisici influenzano l'intricato processo dell'ereditarietà. Le sostanze chimiche genotossiche sono definite come composti in grado di modificare il materiale ereditario delle cellule viventi. La probabilità che una particolare sostanza chimica causi un danno genetico dipende inevitabilmente da diverse variabili, tra cui il livello di esposizione dell'organismo alla sostanza chimica, la distribuzione e la ritenzione della sostanza chimica una volta entrata nell'organismo, l'efficienza dell'attivazione metabolica e/o dei sistemi di disintossicazione in tessuti bersaglio e la reattività della sostanza chimica o dei suoi metaboliti con le macromolecole critiche all'interno delle cellule. La probabilità che il danno genetico causi la malattia dipende in ultima analisi dalla natura del danno, dalla capacità della cellula di riparare o amplificare il danno genetico, dall'opportunità di esprimere qualunque alterazione sia stata indotta e dalla capacità del corpo di riconoscere e sopprimere la moltiplicazione di cellule aberranti.
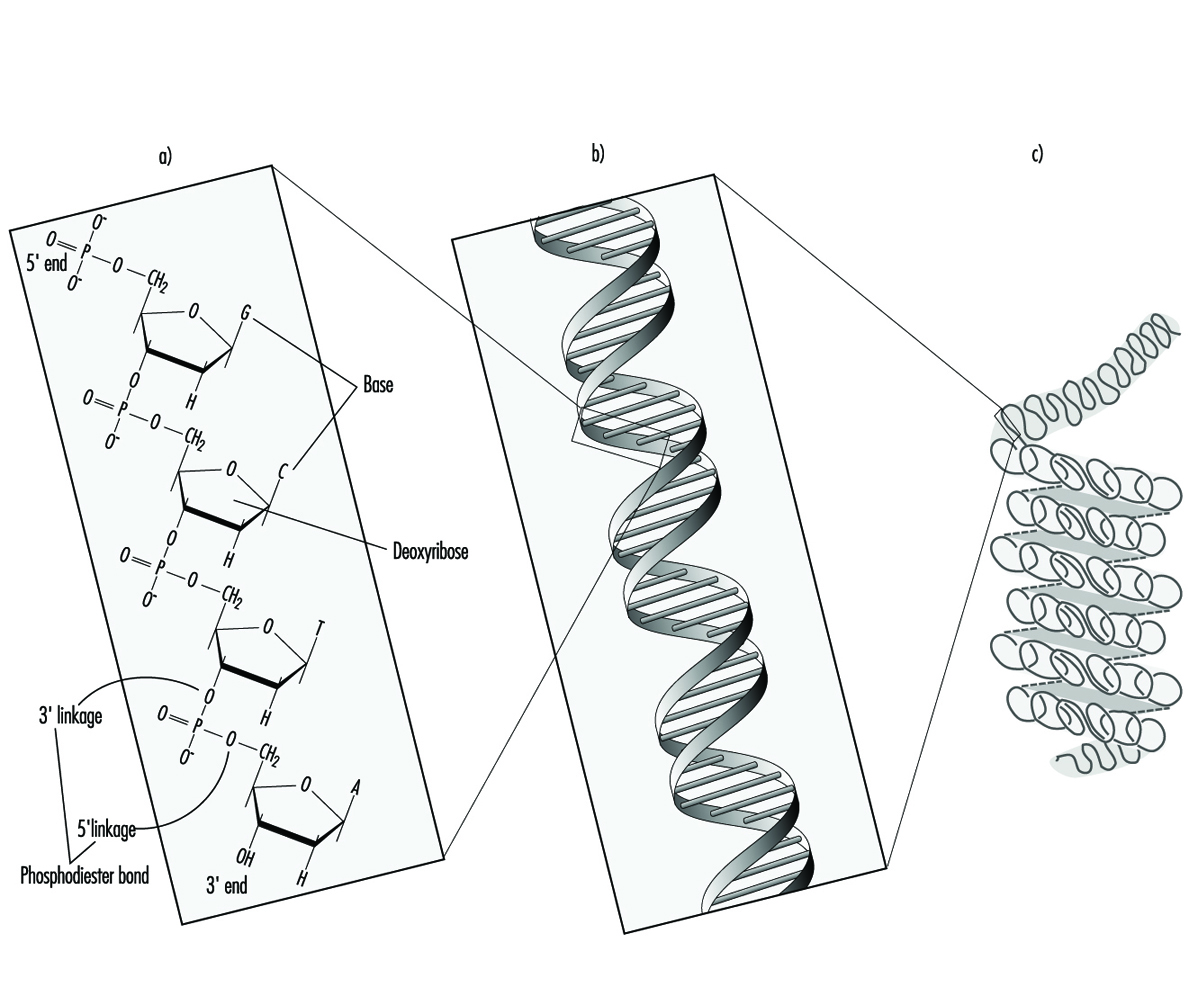
Negli organismi superiori, le informazioni ereditarie sono organizzate nei cromosomi. I cromosomi sono costituiti da filamenti strettamente condensati di DNA associato a proteine. All'interno di un singolo cromosoma, ogni molecola di DNA esiste come una coppia di lunghe catene non ramificate di subunità nucleotidiche collegate tra loro da legami fosfodiestere che uniscono il carbonio 5 di una porzione di desossiribosio al carbonio 3 della successiva (figura 1). Inoltre, una delle quattro diverse basi nucleotidiche (adenina, citosina, guanina o timina) è attaccata a ciascuna subunità di desossiribosio come perline su un filo. Tridimensionalmente, ogni coppia di filamenti di DNA forma una doppia elica con tutte le basi orientate verso l'interno della spirale. All'interno dell'elica, ogni base è associata alla sua base complementare sul filamento di DNA opposto; il legame idrogeno determina un accoppiamento forte e non covalente di adenina con timina e guanina con citosina (figura 1). Poiché la sequenza delle basi nucleotidiche è complementare per l'intera lunghezza della molecola di DNA duplex, entrambi i filamenti portano essenzialmente la stessa informazione genetica. Infatti, durante la replicazione del DNA ogni filamento funge da stampo per la produzione di un nuovo filamento partner.
Figura 1. L'organizzazione (a) primaria, (b) secondaria e (c) terziaria delle informazioni ereditarie umane
 Utilizzando l'RNA e una serie di diverse proteine, la cellula alla fine decifra le informazioni codificate dalla sequenza lineare di basi all'interno di regioni specifiche del DNA (geni) e produce proteine essenziali per la sopravvivenza cellulare di base, nonché per la normale crescita e differenziazione. In sostanza, i nucleotidi funzionano come un alfabeto biologico utilizzato per codificare gli amminoacidi, i mattoni delle proteine.
Utilizzando l'RNA e una serie di diverse proteine, la cellula alla fine decifra le informazioni codificate dalla sequenza lineare di basi all'interno di regioni specifiche del DNA (geni) e produce proteine essenziali per la sopravvivenza cellulare di base, nonché per la normale crescita e differenziazione. In sostanza, i nucleotidi funzionano come un alfabeto biologico utilizzato per codificare gli amminoacidi, i mattoni delle proteine.
Quando vengono inseriti nucleotidi errati o i nucleotidi vengono persi, o quando vengono aggiunti nucleotidi non necessari durante la sintesi del DNA, l'errore viene chiamato mutazione. È stato stimato che si verifica meno di una mutazione ogni 109 nucleotidi incorporati durante la normale replicazione delle cellule. Sebbene le mutazioni non siano necessariamente dannose, le alterazioni che causano l'inattivazione o la sovraespressione di geni importanti possono provocare una varietà di disturbi, tra cui cancro, malattie ereditarie, anomalie dello sviluppo, infertilità e morte embrionale o perinatale. Molto raramente, una mutazione può portare a una maggiore sopravvivenza; tali occorrenze sono la base della selezione naturale.
Sebbene alcune sostanze chimiche reagiscano direttamente con il DNA, la maggior parte richiede l'attivazione metabolica. In quest'ultimo caso, gli intermedi elettrofili come gli epossidi o gli ioni di carbonio sono in ultima analisi responsabili dell'induzione di lesioni in una varietà di siti nucleofili all'interno del materiale genetico (figura 2). In altri casi, la genotossicità è mediata da sottoprodotti dell'interazione dei composti con lipidi intracellulari, proteine o ossigeno.
Figura 2. Bioattivazione di: a) benzo(a)pirene; e b) N-nitrosodimetilammina

A causa della loro relativa abbondanza nelle cellule, le proteine sono il bersaglio più frequente dell'interazione tossica. Tuttavia, la modifica del DNA è di maggiore preoccupazione a causa del ruolo centrale di questa molecola nella regolazione della crescita e della differenziazione attraverso più generazioni di cellule.
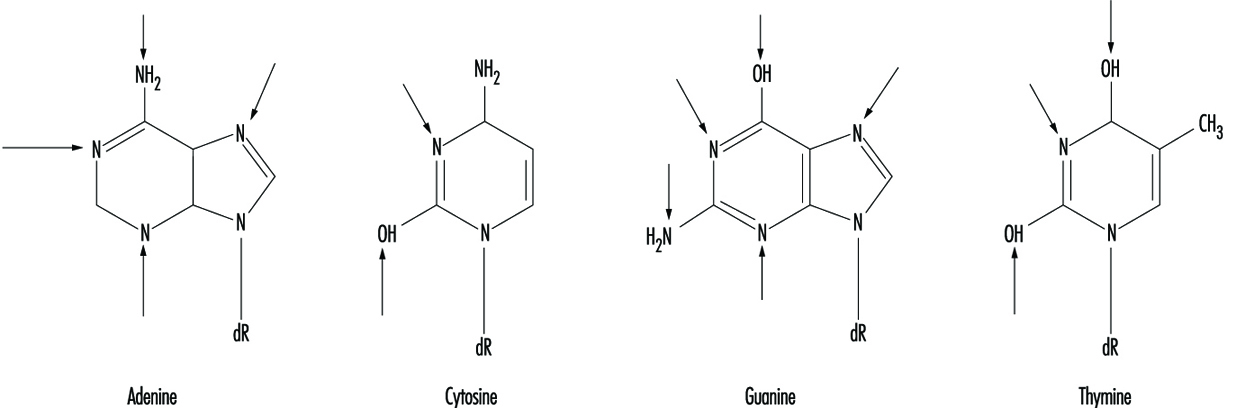
A livello molecolare, i composti elettrofili tendono ad attaccare l'ossigeno e l'azoto nel DNA. I siti che sono più inclini alla modifica sono illustrati nella figura 3. Sebbene gli ossigeni all'interno dei gruppi fosfato nella spina dorsale del DNA siano anche bersagli per la modificazione chimica, si ritiene che il danno alle basi sia biologicamente più rilevante poiché questi gruppi sono considerati i principali elementi nella molecola del DNA.
Figura 3. Siti primari di danno al DNA indotto chimicamente
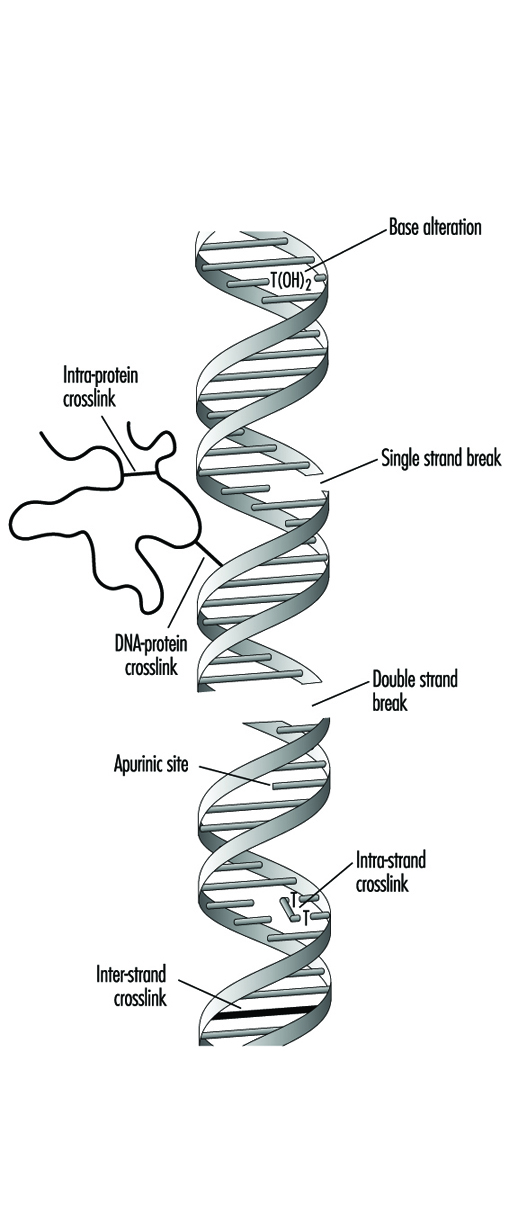
I composti che contengono una porzione elettrofila tipicamente esercitano genotossicità producendo mono-addotti nel DNA. Allo stesso modo, i composti che contengono due o più frazioni reattive possono reagire con due diversi centri nucleofili e quindi produrre legami incrociati intra o intermolecolari nel materiale genetico (figura 4). I legami incrociati tra DNA-DNA e DNA-proteina possono essere particolarmente citotossici poiché possono formare blocchi completi alla replicazione del DNA. Per ovvie ragioni, la morte di una cellula elimina la possibilità che venga mutata o trasformata neoplasticamente. Gli agenti genotossici possono anche agire inducendo rotture nello scheletro del fosfodiestere o tra basi e zuccheri (producendo siti abasici) nel DNA. Tali rotture possono essere un risultato diretto della reattività chimica nel sito del danno o possono verificarsi durante la riparazione di uno dei suddetti tipi di lesione del DNA.
Figura 4. Vari tipi di danno al complesso proteina-DNA
Negli ultimi trenta o quarant'anni sono state sviluppate diverse tecniche per monitorare il tipo di danno genetico indotto da varie sostanze chimiche. Tali saggi sono descritti in dettaglio altrove in questo capitolo e Enciclopedia.
L'errata replicazione di "microlesioni" come mono-addotti, siti abasici o rotture a singolo filamento può in ultima analisi provocare sostituzioni di coppie di basi nucleotidiche o l'inserimento o la delezione di brevi frammenti polinucleotidici nel DNA cromosomico. Al contrario, le "macrolesioni", come addotti voluminosi, collegamenti incrociati o rotture a doppio filamento possono innescare l'acquisizione, la perdita o il riarrangiamento di pezzi relativamente grandi di cromosomi. In ogni caso, le conseguenze possono essere devastanti per l'organismo poiché ognuno di questi eventi può portare alla morte cellulare, alla perdita di funzione o alla trasformazione maligna delle cellule. Il modo esatto in cui il danno al DNA provoca il cancro è in gran parte sconosciuto. Attualmente si ritiene che il processo possa comportare un'attivazione inappropriata di proto-oncogeni come il mio c ed ras, e/o inattivazione di geni soppressori tumorali recentemente identificati come p53. L'espressione anormale di entrambi i tipi di geni abroga i normali meccanismi cellulari per controllare la proliferazione e/o la differenziazione cellulare.
La preponderanza di prove sperimentali indica che lo sviluppo del cancro in seguito all'esposizione a composti elettrofili è un evento relativamente raro. Ciò può essere spiegato, in parte, dalla capacità intrinseca della cellula di riconoscere e riparare il DNA danneggiato o dall'incapacità delle cellule con DNA danneggiato di sopravvivere. Durante la riparazione, la base danneggiata, il nucleotide o il breve tratto di nucleotidi che circonda il sito del danno viene rimosso e (usando il filamento opposto come modello) viene sintetizzato e inserito in posizione un nuovo pezzo di DNA. Per essere efficace, la riparazione del DNA deve avvenire con grande accuratezza prima della divisione cellulare, prima delle opportunità di propagazione della mutazione.
Studi clinici hanno dimostrato che le persone con difetti ereditari nella capacità di riparare il DNA danneggiato spesso sviluppano tumori e/o anomalie dello sviluppo in tenera età (tabella 1). Tali esempi forniscono una forte evidenza che collega l'accumulo di danni al DNA alla malattia umana. Allo stesso modo, gli agenti che promuovono la proliferazione cellulare (come il tetradecanoilforbolo acetato) spesso aumentano la carcinogenesi. Per questi composti, l'aumentata probabilità di trasformazione neoplastica può essere una diretta conseguenza di una diminuzione del tempo a disposizione della cellula per effettuare un'adeguata riparazione del DNA.
Tabella 1. Malattie ereditarie a rischio di cancro che sembrano comportare difetti nella riparazione del DNA
| Sindrome | Sintomi | Fenotipo cellulare |
| Atassia teleangectasia | Deterioramento neurologico immunodeficienza Elevata incidenza di linfomi |
Ipersensibilità alle radiazioni ionizzanti e ad alcuni agenti alchilanti. Replicazione disregolata del DNA danneggiato (può indicare un tempo ridotto per la riparazione del DNA) |
| Sindrome di Bloom | Anomalie dello sviluppo Lesioni sulla pelle esposta Alta incidenza di tumori del sistema immunitario e del tratto gastrointestinale |
Alta frequenza di aberrazioni cromosomiche Legatura difettosa di rotture associate alla riparazione del DNA |
| Anemia di Fanconi | Ritardo della crescita Alta incidenza di leucemia |
Ipersensibilità agli agenti reticolanti Alta frequenza di aberrazioni cromosomiche Riparazione difettosa dei collegamenti incrociati nel DNA |
| Cancro del colon ereditario non poliposico | Alta incidenza di cancro al colon | Difetto nella riparazione del mismatch del DNA (quando si verifica l'inserimento di un nucleotide errato durante la replicazione) |
| Xeroderma pigmentoso | Alta incidenza di epitelioma sulle aree esposte della pelle Compromissione neurologica (in molti casi) |
Ipersensibilità ai raggi UV e a molti agenti cancerogeni chimici Difetti nella riparazione dell'escissione e/o nella replicazione del DNA danneggiato |
Le prime teorie su come le sostanze chimiche interagiscono con il DNA possono essere fatte risalire a studi condotti durante lo sviluppo del gas mostarda per l'uso in guerra. Un'ulteriore comprensione è nata dagli sforzi per identificare agenti antitumorali che arrestassero selettivamente la replicazione delle cellule tumorali in rapida divisione. La crescente preoccupazione del pubblico per i pericoli nel nostro ambiente ha stimolato ulteriori ricerche sui meccanismi e le conseguenze dell'interazione chimica con il materiale genetico. Esempi di vari tipi di sostanze chimiche che esercitano genotossicità sono presentati nella tabella 2.
Tabella 2. Esempi di sostanze chimiche che presentano genotossicità nelle cellule umane
| Classe di sostanza chimica | Esempio | Fonte di esposizione | Probabile lesione genotossica |
| Le aflatossine | Aflatossina B1 | Alimenti contaminati | Addotti voluminosi del DNA |
| Ammine aromatiche | 2-acetilamminofluorene | Ambientali | Addotti voluminosi del DNA |
| Chinoni di aziridina | Mitomicina C | Chemioterapia contro il cancro | Mono-addotti, reticolazioni interfilari e rotture a singolo filamento nel DNA. |
| Idrocarburi clorurati | Cloruro di vinile | Ambientali | Mono-addotti nel DNA |
| Metalli e composti metallici | cisplatino | Chemioterapia contro il cancro | Entrambi i collegamenti incrociati intra e inter filamento nel DNA |
| Composti di nichel | Ambientali | Mono-addotti e rotture a singolo filamento nel DNA | |
| Mostarde di azoto | Ciclofosfamide | Chemioterapia contro il cancro | Mono-addotti e reticolazioni interfilari nel DNA |
| Le nitrosammine | N-nitrosodimetilammina | Alimenti contaminati | Mono-addotti nel DNA |
| Idrocarburi policiclici aromatici | Il benzo (a) pirene | Ambientali | Addotti voluminosi del DNA |