Les cellules nerveuses sont les unités fonctionnelles du système nerveux. On pense que le système nerveux compte dix milliards de ces cellules, appelées neurones ainsi que glie, la glie étant présente en plus grand nombre que les neurones.
Le Neurone
La figure 1 est un schéma idéalisé d'un neurone avec ses trois caractéristiques structurelles les plus importantes : le corps cellulaire, les dendrites et l'axone terminal.
Figure 1. L'anatomie du neurone
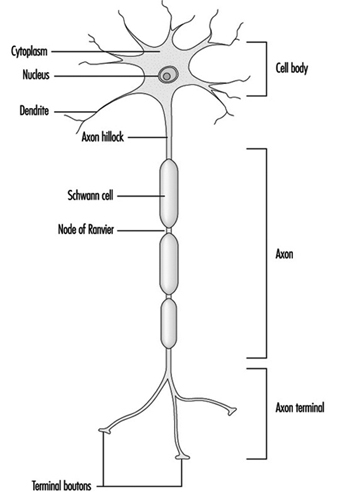
Les dendrites sont des processus finement ramifiés apparaissant près du corps cellulaire d'un neurone. Les dendrites reçoivent des effets excitateurs ou inhibiteurs via des messagers chimiques appelés neurotransmetteurs. Le cytoplasme est le matériau du corps cellulaire dans lequel se trouvent les organites, y compris le noyau cellulaire, et d'autres inclusions Figure 2. Le noyau contient la chromatine ou le matériel génétique de la cellule.
Figure 2. Les organites
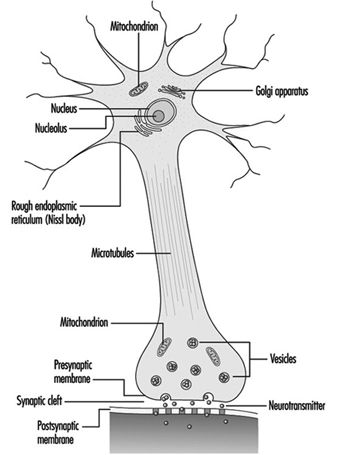
Le noyau de la cellule nerveuse est atypique par rapport à celui des autres cellules vivantes en ce que, bien qu'il contienne le matériel génétique acide désoxyribonucléique (ADN), l'ADN n'intervient pas dans le processus de division cellulaire ; c'est-à-dire qu'après avoir atteint la maturité, les cellules nerveuses ne se divisent pas. (Une exception à cette règle sont les neurones de la muqueuse nasale (épithélium olfactif).) Le noyau est riche en acide ribonucléique (ARN), qui est nécessaire à la synthèse des protéines. Trois types de protéines ont été identifiées : les protéines cytosoliques, qui forment les éléments fibrillaires de la cellule nerveuse ; les protéines intracondriales, qui génèrent de l'énergie pour l'activité cellulaire ; et les protéines qui forment les membranes et les produits de sécrétion. Les neurones sont maintenant conçus comme des cellules sécrétoires modifiées. Des granules sécrétoires sont formés, stockés dans des vésicules synaptiques et ensuite libérés sous forme de substances neurotransmetteurs, les messagers chimiques entre les cellules nerveuses.
Les éléments fibrillaires, qui forment le squelette du neurone, participent à la fonction trophique du neurone, agissant comme véhicules de transmission. Le transport axonal peut être antérograde (du corps cellulaire à l'axone terminal) et rétrograde (de l'axone terminal au corps cellulaire). Du plus épais au plus fin, trois types d'éléments fibrillaires sont reconnus : les microtubules, les neurofilaments et les microfilaments.
Cellules gliales
Contrairement aux neurones, les cellules gliales ne véhiculent pas, par elles-mêmes, de messages électriques. Il existe deux types de cellules gliales : les macroglie et par microglie. La macroglie est un nom donné à au moins trois types de cellules : les astrocytes, les oligodendrocytes et les cellules épendymaires. Les cellules microgliales sont principalement des cellules charognardes pour éliminer les débris après une lésion neurale ou une infection.
Les cellules gliales ont également des caractéristiques microscopiques et ultramicroscopiques distinctives. Les cellules gliales soutiennent physiquement les neurones, mais un certain nombre de propriétés physiologiques commencent également à être comprises. Parmi les interactions neurone-glie les plus importantes figurent le rôle de la cellule gliale dans l'apport de nutriments aux neurones, l'élimination des fragments de neurones après leur mort et, surtout, la contribution au processus de communication chimique. Contrairement aux neurones, les cellules gliales peuvent se diviser et ainsi se reproduire. Les tumeurs du système nerveux, par exemple, résultent d'une reproduction anormale des cellules gliales.
Myéline
Ce qui apparaît dans l'observation macroscopique du tissu neural comme « matière grise » et « matière blanche » a une base microscopique et biochimique. Au microscope, la matière grise contient les corps cellulaires neuronaux, tandis que la matière blanche est l'endroit où se trouvent les fibres neurales ou les axones. L'aspect « blanc » est dû à une gaine - composée d'un corps gras appelé myéline - recouvrant ces fibres. La myéline des nerfs périphériques provient de la membrane de la cellule de Schwann qui s'enroule autour de l'axone. La myéline des fibres du système nerveux central est fournie par les membranes des oligodendrocytes (une variété de cellules gliales). Les oligodendrocytes myélinisent généralement plusieurs axones, alors que la cellule de Schwann est associée à un seul axone. Une discontinuité de la gaine de myéline - désignée sous le nom de nœuds de Ranvier - existe entre les cellules de Schwann continues ou les oligodendrocytes. On estime que dans la voie motrice centrale la plus longue, jusqu'à 2,000 XNUMX cellules de Schwann forment la couverture de myéline. La myéline, dont le rôle est de faciliter la propagation du potentiel d'action, peut être une cible spécifique des agents neurotoxiques. Une classification morphologique des substances neurotoxiques décrit les modifications neuropathologiques caractéristiques de la myéline comme des myélinopathies.
Fonction trophique du neurone
Les fonctions normales du neurone comprennent la synthèse des protéines, le transport axonal, la génération et la conduction du potentiel d'action, la transmission synaptique, ainsi que la formation et le maintien de la myéline. Certaines des fonctions trophiques fondamentales du neurone ont été décrites dès le 19ème siècle en sectionnant les axones (axotomie). Parmi les processus découverts, l'un des plus importants était la dégénérescence wallérienne - d'après Waller, le physiologiste anglais qui l'a décrite.
La dégénérescence wallérienne offre une bonne occasion de décrire des changements bien connus dans les organites à la suite de dommages traumatiques ou toxiques. Entre parenthèses, les termes utilisés pour décrire la dégénérescence wallérienne produite par l'axotomie traumatique sont les mêmes que ceux utilisés pour décrire les changements résultant d'agents neurotoxiques. Au niveau cellulaire, les changements neuropathologiques résultant de dommages toxiques au tissu neural sont beaucoup plus complexes que ceux qui se produisent à la suite de dommages traumatiques. Ce n'est que récemment que des changements dans les neurones affectés par des agents neurotoxiques ont été observés.
Vingt-quatre heures après la coupe de l'axone, la caractéristique la plus distinctive est le gonflement des deux côtés du traumatisme mécanique. Le gonflement résulte de l'accumulation de fluides et d'éléments membraneux des deux côtés du site de la blessure. Ces changements ne sont pas sans rappeler ceux observés dans une route à double sens inondée par la pluie avec des véhicules arrêtés des deux côtés de la zone inondée. Dans cette analogie, les véhicules en panne sont le gonflement. Après quelques jours, la régénération des axones engainés, c'est-à-dire ceux recouverts de myéline, se produit. Les germes poussent à partir du moignon proximal en se déplaçant au rythme de 1 à 3 mm par jour. Dans des conditions favorables, les germes atteignent la souche distale (plus éloignée du corps cellulaire). Lorsque la rénervation - la jonction des moignons - est terminée, les caractéristiques de base de la transmission normale ont été rétablies. Le corps cellulaire du neurone lésé subit de profonds changements structurels dans la synthèse des protéines et le transport axonal.
Si la neurobiologie moléculaire est dite une discipline jeune, la neurobiologie des processus neurotoxiques est encore plus jeune, et encore balbutiante. Certes, la base moléculaire de l'action de nombreuses neurotoxines et agents pharmacologiques est maintenant bien comprise. Mais à quelques exceptions notables près (p. ex. plomb, méthylmercure, acrylamide), la base moléculaire de la toxicité de la grande majorité des agents environnementaux et neurotoxiques est inconnue. C'est pourquoi, au lieu de décrire la neurobiologie moléculaire d'un groupe restreint d'agents neurotoxiques professionnels et environnementaux, nous sommes toujours obligés de nous référer aux stratégies et exemples relativement abondants de la neuropharmacologie classique ou des travaux de fabrication de médicaments modernes.
Neurotransmetteurs
Un neurotransmetteur est une substance chimique qui, lorsqu'elle est libérée des terminaisons axonales par le potentiel d'action, produit le changement momentané de potentiel électrique lorsqu'une autre fibre nerveuse est stimulée. Les neurotransmetteurs stimulent ou inhibent les neurones adjacents ou les organes effecteurs tels que les muscles et les glandes. Les neurotransmetteurs connus et leurs voies neuronales font maintenant l'objet d'études intensives et de nouveaux sont constamment découverts. On comprend maintenant que certains troubles neurologiques et psychiatriques sont causés par des modifications chimiques de la neurotransmission, par exemple la myasthénie grave, la maladie de Parkinson, certaines formes de troubles affectifs tels que la dépression, une distorsion grave des processus de pensée comme dans la schizophrénie et la maladie d'Alzheimer. Bien que d'excellents rapports isolés sur l'effet de plusieurs agents neurotoxiques environnementaux et professionnels sur la neurotransmission aient été publiés, le corpus de connaissances est maigre par rapport à celui existant pour les maladies neuropsychiatriques. Les études pharmacologiques des médicaments fabriqués nécessitent une compréhension de la façon dont les médicaments affectent la neurotransmission. La fabrication de médicaments et la recherche sur la neurotransmission sont donc intimement liées. L'évolution des points de vue sur l'action des médicaments a été résumée par Feldman et Quenzer (1984).
Les effets des agents neurotoxiques sur la neurotransmission sont caractérisés par l'endroit du système nerveux où ils agissent, leurs récepteurs chimiques, l'évolution temporelle de leurs effets, si les agents neurotoxiques facilitent, bloquent ou inhibent la neurotransmission, ou si les agents neurotoxiques modifient la terminaison ou l'élimination de la l'action pharmacologique du neurotransmetteur.
Une difficulté rencontrée par les neuroscientifiques est la nécessité de lier les processus connus qui se produisent au niveau moléculaire dans le neurone avec des événements au niveau cellulaire, qui à leur tour peuvent expliquer comment les changements neuropsychologiques normaux et pathologiques se produisent, comme indiqué clairement dans ce qui suit qui à un s'applique toujours dans une large mesure : « (A)t au niveau moléculaire, une explication de l'action d'un médicament est souvent possible ; au niveau cellulaire, une explication est parfois possible, mais au niveau comportemental, notre ignorance est abyssale » (Cooper, Bloom et Roth 1986).
Les principaux composants du système nerveux
La connaissance des principaux composants du système nerveux est essentielle pour la compréhension des manifestations neuropsychologiques globales des maladies neurotoxiques, la justification de l'utilisation de techniques spécifiques pour l'évaluation des fonctions du système nerveux et la compréhension des mécanismes pharmacologiques de l'action neurotoxique. D'un point de vue fonctionnel, le système nerveux peut être divisé en deux grands compartiments : système nerveux somatique transmet des informations sensorielles (toucher, température, douleur et position des membres - même lorsque les yeux sont fermés) des segments du corps et transporte les voies neuronales qui innervent et contrôlent le mouvement des muscles squelettiques, tels que ceux des bras, des doigts, des jambes et orteils. Le système nerveux viscéral contrôle les organes internes qui ne sont normalement pas sous l'influence des vaisseaux sanguins, la dilatation et la constriction des pupilles des yeux, etc.
D'un point de vue anatomique, quatre composantes principales doivent être identifiées : la le système nerveux central, système nerveux périphérique y compris les nerfs crâniens, le système autonome et par système neuroendocrinien.
Le système nerveux central
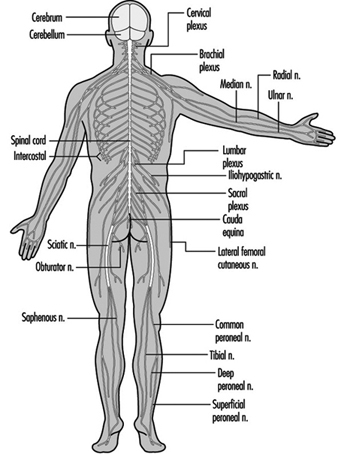
Le système nerveux central contient le cerveau et la moelle épinière Figure 3. Le cerveau se trouve dans la cavité crânienne et est protégé par les méninges. Il est divisé en trois composantes principales; dans l'ordre croissant - c'est-à-dire de la partie caudale (queue) à la partie cervicale (tête) du système nerveux - ce sont le cerveau postérieur (également appelé rhombencéphale), le cerveau moyen (le mésencéphale) et le cerveau antérieur (le proscencéphale).
Figure 3. Les divisions centrale et périphérique du système nerveux
Le cerveau postérieur
Les trois principaux composants du cerveau postérieur sont le bulbe rachidien, le pont et le cervelet figure 4.
Figure 4. Le cerveau vu de profil.
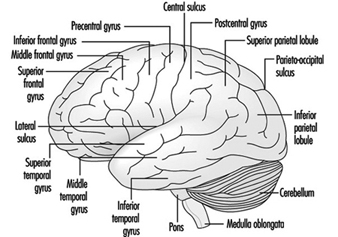
Le bulbe rachidien contient des structures neurales qui contrôlent la fréquence cardiaque et la respiration, parfois les cibles d'agents neurotoxiques et de médicaments causant la mort. Situé entre le bulbe rachidien et le mésencéphale, le pons (pont) tire son nom du grand nombre de fibres traversant sa face antérieure en route vers les hémisphères cérébelleux. Le cervelet - en latin, petit cerveau - a une apparence typiquement ondulée. Le cervelet reçoit des informations sensorielles et envoie des messages moteurs essentiels à la coordination motrice. Il est responsable (entre autres fonctions) de l'exécution des mouvements fins. Cette planification - ou programmation - nécessite un timing adéquat des entrées sensorielles et des réponses motrices. Le cervelet est souvent la cible de nombreux agents neurotoxiques, par exemple les boissons alcoolisées, de nombreux solvants industriels, le plomb, qui affectent les réponses motrices.
Le mésencéphale
Le mésencéphale est une partie étroite du cerveau reliant le cerveau postérieur au cerveau antérieur. Les structures du mésencéphale sont l'aqueduc cérébral, le tectum, les pédoncules cérébraux, la substantia nigra et le noyau rouge. L'aqueduc cérébral est un canal qui relie le troisième au quatrième ventricule (cavités remplies de liquide du cerveau); le liquide céphalo-rachidien (LCR) s'écoule par cette ouverture.
Le cerveau antérieur
Cette partie du cerveau est subdivisée en diencéphale (« entre le cerveau ») et le cerveau. Les principales régions du diencéphale sont le thalamus et l'hypothalamus. « Thalamus » signifie « chambre intérieure ». Les thalami sont constitués de groupements neuronaux, appelés noyaux, qui ont cinq fonctions principales :
- recevoir des informations sensorielles et les envoyer aux zones primaires du cortex cérébral
- envoyer des informations sur les mouvements en cours aux zones motrices du cortex cérébral
- envoyer des informations sur l'activité du système limbique aux zones du cortex cérébral liées à ce système
- envoyer des informations sur l'activité intrathalamique aux zones d'association du cortex cérébral
- envoyer des informations sur l'activité de formation réticulaire du tronc cérébral à des zones étendues du cortex cérébral.
Le nom hypothalamus signifie « sous le thalamus ». Il forme la base du troisième ventricule, un point de référence important pour l'imagerie du cerveau. L'hypothalamus est une structure neuronale complexe et minuscule responsable de nombreux aspects du comportement tels que les pulsions biologiques de base, la motivation et l'émotion. C'est le lien entre le système nerveux et le système neuroendocrinien, dont il sera question ci-dessous. L'hypophyse (également appelée hypophyse) est reliée par des neurones aux noyaux hypothalamiques. Il est bien établi que les cellules nerveuses hypothalamiques remplissent de nombreuses fonctions neurosécrétoires. L'hypothalamus est lié à de nombreuses autres régions majeures du cerveau, notamment le rhinencéphale - le cortex primitif initialement associé à l'olfaction - et le système limbique, y compris l'hippocampe.
Le cortex cérébral est le plus grand composant du cerveau, composé de deux hémisphères cérébraux reliés par une masse de matière blanche appelée corps calleux. Le cortex cérébral est la couche superficielle de chaque hémisphère cérébral. Les sillons profonds du cortex cérébral (les sillons central et latéral Figure 4) sont pris comme points de référence pour séparer les régions anatomiques du cerveau. Le lobe frontal se trouve devant le sillon central. Le lobe pariétal commence à l'arrière du sillon central et se trouve à côté du lobe occipital, qui occupe la partie postérieure du cerveau. Le lobe temporal commence bien à l'intérieur du repliement du sillon latéral et s'étend dans les aspects ventraux des hémisphères cérébraux. Deux composants importants du cerveau sont les ganglions de la base et le système limbique.
Les ganglions de la base sont des noyaux, c'est-à-dire des amas de cellules nerveuses, situés vers le centre du cerveau. Les noyaux gris centraux comprennent les principaux centres du système moteur extra-pyramidal. (Le système pyramidal, auquel le terme est opposé, est impliqué dans le contrôle volontaire du mouvement.) Le système extrapyramidal est sélectivement affecté par de nombreux agents neurotoxiques (par exemple, le manganèse). Au cours des deux dernières décennies, d'importantes découvertes ont été faites concernant le rôle que ces noyaux jouent dans plusieurs maladies neurales dégénératives (par exemple, la maladie de Parkinson, la chorée de Huntington).
Le système limbique est composé de structures neuronales alambiquées se ramifiant dans de nombreuses directions et établissant des connexions avec de nombreuses « anciennes » régions du cerveau, en particulier avec l'hypothalamus. Il est impliqué dans le contrôle de l'expression émotionnelle. On pense que l'hippocampe est une structure où se produisent de nombreux processus de mémoire.
La moelle épinière
La moelle épinière est une structure blanchâtre située dans le canal vertébral. Elle est divisée en quatre régions : cervicale, thoracique, lombaire et sacro-coccyxienne. Les deux caractéristiques les plus facilement reconnaissables de la moelle épinière sont la substance grise contenant les corps cellulaires des neurones et la substance blanche contenant les axones myélinisés des neurones. La région ventrale de la matière grise de la moelle épinière contient des cellules nerveuses qui régulent la fonction motrice ; la région médiane de la moelle épinière thoracique est associée aux fonctions autonomes. La partie dorsale reçoit les informations sensorielles des nerfs rachidiens.
Le système nerveux périphérique
Le système nerveux périphérique comprend les neurones situés à l'extérieur du système nerveux central. Le terme périphérique décrit la distribution anatomique de ce système, mais fonctionnellement il est artificiel. Les corps cellulaires des fibres motrices périphériques, par exemple, sont situés dans le système nerveux central. En neurotoxicologie expérimentale, clinique et épidémiologique, le terme système nerveux périphérique (PNS) décrit un système sélectivement vulnérable aux effets des agents toxiques et capable de se régénérer.
Les nerfs spinaux
Les racines ventrales et dorsales sont l'endroit où les nerfs périphériques entrent et sortent de la moelle épinière sur toute sa longueur. Les vertèbres adjacentes contiennent des ouvertures pour permettre aux fibres racinaires formant les nerfs rachidiens de quitter le canal rachidien. Il existe 31 paires de nerfs rachidiens, nommés selon la région de la colonne vertébrale à laquelle ils sont associés : 8 cervicaux, 12 thoraciques, 5 lombaires, 5 sacrés et 1 coccyxien. Un métamère est une région du corps innervée par un nerf spinal figure 5.
Figure 5. La distribution segmentaire des nerfs rachidiens (les métamères).
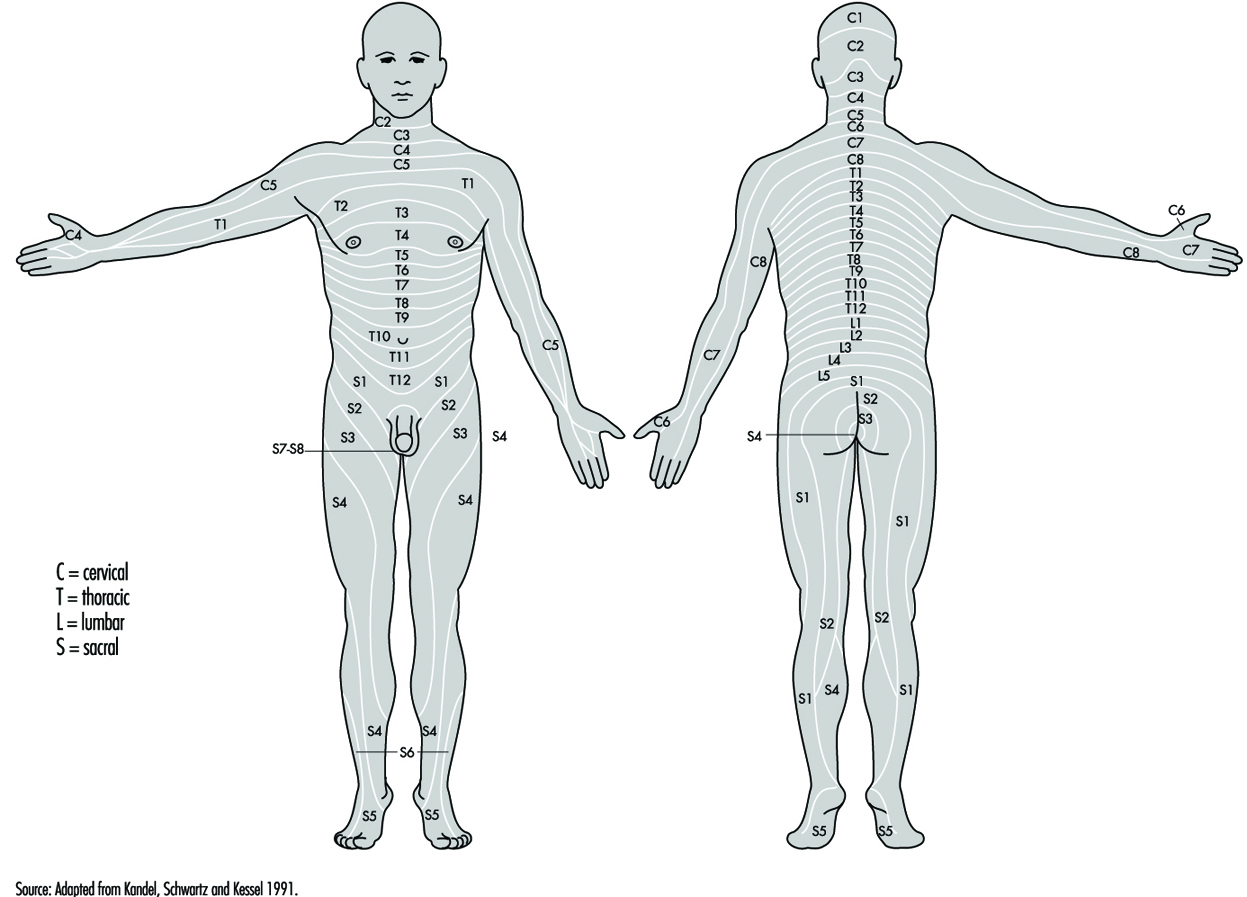
En examinant attentivement les fonctions motrices et sensorielles des métamères, les neurologues peuvent déduire l'emplacement des lésions où des dommages se sont produits.
Tableau 1. Noms et fonctions principales de chaque paire de nerfs crâniens
| Nerf1 | Conduit les impulsions | Les fonctions |
| I. Olfactif | Du nez au cerveau | Odorat |
| II. Optique | De l'œil au cerveau | Vision |
| III. Oculomoteur | Du cerveau aux muscles oculaires | Mouvements oculaires |
| IV. Trochléaire | Du cerveau aux muscles oculaires externes | Mouvements oculaires |
| V. Trijumeau (ou trifacial) |
De la peau et des muqueuses de la tête et des dents au cerveau ; aussi du cerveau aux muscles masticateurs | Sensations du visage, du cuir chevelu et des dents ; mouvements de mastication |
| VI. abducens | Du cerveau aux muscles oculaires externes | Tourner les yeux vers l'extérieur |
| VII. Soin du visage | Des papilles gustatives de la langue au cerveau ; du cerveau aux muscles du visage | Sens du goût; contraction des muscles de l'expression faciale |
| VIII. Acoustique | De l'oreille au cerveau | Audience; sens de l'équilibre |
| IX. Glossopharyngien | De la gorge et des papilles gustatives de la langue au cerveau ; également du cerveau aux muscles de la gorge et aux glandes salivaires | Sensations de gorge, goût, mouvements de déglutition, sécrétion de salive |
| X. Vague | De la gorge, du larynx et des organes des cavités thoracique et abdominale au cerveau ; également du cerveau aux muscles de la gorge et aux organes des cavités thoracique et abdominale | Sensations de la gorge, du larynx et des organes thoraciques et abdominaux ; déglutition, production de voix, ralentissement du rythme cardiaque, accélération du péristaltisme |
| XI. Accessoire rachidien | Du cerveau à certains muscles des épaules et du cou | Mouvements des épaules ; mouvements tournants de la tête |
| XII. Hypoglosse | Du cerveau aux muscles de la langue | Mouvements de la langue |
1 La première lettre des mots de la phrase suivante sont les premières lettres des noms des nerfs crâniens : "On Old Olympus' Tiny Tops A Finland and German Viewed Some Hops". De nombreuses générations d'étudiants ont utilisé cette phrase ou une phrase similaire pour les aider à se souvenir des noms des nerfs crâniens.
Les nerfs crâniens
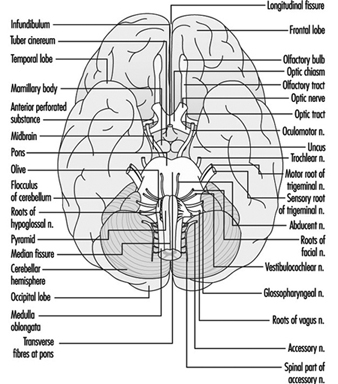
Tige du cerveau est un terme global qui désigne la région du système nerveux qui comprend la moelle, le pont et le mésencéphale. Le tronc cérébral est une continuation de la moelle épinière vers le haut et vers l'avant (ventralement). C'est dans cette région que la plupart des nerfs crâniens font leurs sorties et leurs entrées. Il y a 12 paires de nerfs crâniens ; Le tableau 1 décrit le nom et la fonction principale de chaque paire et la figure 6 montre l'entrée et la sortie de certains nerfs crâniens dans le cerveau.
Figure 6. Le cerveau vu de dessous avec l'entrée et la sortie de nombreux nerfs crâniens.
Le système nerveux autonome
Le système nerveux autonome est la partie du système nerveux contrôlant l'activité des composants viscéraux du corps humain. Il est dit « autonome » car il remplit ses fonctions automatiquement, ce qui signifie que son fonctionnement ne peut pas être facilement contrôlé à volonté. D'un point de vue anatomique, le système autonome a deux composants principaux : le système nerveux sympathique et le système nerveux parasympathique. Les nerfs sympathiques contrôlant l'activité viscérale proviennent des parties thoracique et lombaire de la moelle épinière ; les nerfs parasympathiques proviennent du tronc cérébral et de la partie sacrée de la moelle épinière.
D'un point de vue physiologique, aucune généralisation ne peut être faite qui s'applique à la manière dont les systèmes nerveux sympathique et parasympathique contrôlent différents organes du corps. Dans la plupart des cas, les organes viscéraux sont innervés par les deux systèmes, et chaque type a un effet opposé dans un système de freins et contrepoids. Le cœur, par exemple, est innervé par des nerfs sympathiques dont l'excitation produit une accélération du rythme cardiaque, et par des nerfs parasympathiques dont l'excitation produit un ralentissement du rythme cardiaque. L'un ou l'autre système peut stimuler ou inhiber les organes qu'il innerve. Dans d'autres cas, les organes sont majoritairement ou exclusivement contrôlés par l'un ou l'autre système. Une fonction vitale du système nerveux autonome est le maintien de l'homéostasie (état d'équilibre stable) et l'adaptation du corps animal à son environnement extérieur. L'homéostasie est l'état d'équilibre des fonctions corporelles atteint par un processus actif ; le contrôle de la température corporelle, de l'eau et des électrolytes sont tous des exemples de processus homéostatiques.
Du point de vue pharmacologique, il n'y a pas de neurotransmetteur unique associé aux fonctions sympathiques ou parasympathiques, comme on le croyait autrefois. L'ancienne opinion selon laquelle l'acétylcholine était le transmetteur prédominant du système autonome a dû être abandonnée lorsque de nouvelles classes de neurotransmetteurs et de neuromodulateurs ont été découvertes (par exemple, la dopamine, la sérotonine, les purines et divers neuropeptides).
Les neuroscientifiques ont récemment relancé le point de vue comportemental du système nerveux autonome. Le système nerveux autonome est impliqué dans la réaction instinctive de combat ou de fuite encore présente chez l'homme, qui est, pour l'essentiel, à la base des réactions physiologiques provoquées par le stress. Les interactions entre le système nerveux et les fonctions immunologiques sont possibles via le système nerveux autonome. Les émotions qui proviennent du système nerveux autonome peuvent être exprimées via les muscles squelettiques.
Le contrôle autonome des muscles lisses
Les muscles des viscères, à l'exception du cœur, sont les muscles lisses. Le muscle cardiaque a des caractéristiques à la fois du muscle squelettique et du muscle lisse. Comme les muscles squelettiques, les muscles lisses contiennent également les deux protéines actine et, dans de moindres proportions, la myosine. Contrairement aux muscles squelettiques, ils ne présentent pas l'organisation régulière des sarcolèmes, l'unité contractile de la fibre musculaire. Le cœur est unique en ce sens qu'il peut générer une activité myogénique - même après que ses innervations neurales ont été sectionnées, il peut se contracter et se détendre pendant plusieurs heures par lui-même.
Le couplage neuromusculaire des muscles lisses diffère de celui des muscles squelettiques. Dans les muscles squelettiques, la jonction neuromusculaire est le lien entre le nerf et les fibres musculaires. Dans le muscle lisse, il n'y a pas de jonction neuromusculaire ; les terminaisons nerveuses pénètrent dans le muscle, se propageant dans toutes les directions. Les événements électriques à l'intérieur du muscle lisse sont donc beaucoup plus lents que ceux des muscles squelettiques. Enfin, le muscle lisse a la caractéristique unique de présenter des contractions spontanées, telles que celles présentées par l'intestin. Dans une large mesure, le système nerveux autonome régule l'activité spontanée des muscles lisses.
Les composants centraux du système nerveux autonome
Le rôle principal du système nerveux autonome est de réguler l'activité des muscles lisses, du cœur, des glandes du tube digestif, des glandes sudoripares et des glandes surrénales et autres glandes endocrines. Le système nerveux autonome a un composant central - l'hypothalamus, situé à la base du cerveau - où de nombreuses fonctions autonomes sont intégrées. Plus important encore, les composants centraux du système autonome sont directement impliqués dans la régulation des pulsions biologiques (régulation de la température, faim, soif, sexe, miction, défécation, etc.), la motivation, l'émotion et dans une large mesure dans les fonctions "psychologiques". comme les humeurs, les affects et les sentiments.
Système neuroendocrinien
Les glandes sont les organes du système endocrinien. On les appelle glandes endocrines car leurs messages chimiques sont délivrés à l'intérieur du corps, directement dans la circulation sanguine (contrairement aux glandes exocrines, comme les glandes sudoripares, dont les sécrétions apparaissent sur la surface externe du corps). Le système endocrinien assure un contrôle lent mais durable des organes et des tissus par le biais de messagers chimiques appelés hormones. Les hormones sont les principaux régulateurs du métabolisme de l'organisme. Mais, en raison des liens intimes entre les systèmes nerveux central, périphérique et autonome, le système neuroendocrinien- un terme qui capture des liens aussi complexes - est maintenant conçu comme un puissant modificateur de la structure et de la fonction du corps humain et du comportement.
Les hormones ont été définies comme des messagers chimiques qui sont libérés des cellules dans la circulation sanguine pour exercer leur action sur les cellules cibles à une certaine distance. Jusqu'à récemment, les hormones étaient distinguées des neurotransmetteurs, discutés ci-dessus. Ces derniers sont des messagers chimiques libérés des neurones vers une synapse entre les terminaisons nerveuses et un autre neurone ou un effecteur (c'est-à-dire un muscle ou une glande). Cependant, avec la découverte que les neurotransmetteurs classiques tels que la dopamine peuvent également agir comme des hormones, la distinction entre les neurotransmetteurs et les hormones est désormais de moins en moins claire. Ainsi, sur la base de considérations purement anatomiques, les hormones dérivées des cellules nerveuses peuvent être appelées neurohormones. D'un point de vue fonctionnel, le système nerveux peut être considéré comme un véritable système neurosécrétoire.
L'hypothalamus contrôle les fonctions endocriniennes par le biais d'un lien avec la glande pituitaire (également appelée hypophyse, une minuscule glande située à la base du cerveau). Jusqu'au milieu des années 1950, les glandes endocrines étaient considérées comme un système distinct régi par l'hypophyse, souvent appelée «glande maîtresse». A cette époque, une hypothèse neurovasculaire a été avancée qui établissait le rôle fonctionnel des facteurs hypothalamiques/hypophysaires dans le contrôle de la fonction endocrinienne. De ce point de vue, l'hypothalamus endocrinien fournit la dernière voie neuroendocrinienne commune dans le contrôle du système endocrinien. Il est désormais bien établi que le système endocrinien est lui-même régulé par le système nerveux central ainsi que par les apports endocriniens. Ainsi, neuroendocrinologie est maintenant un terme approprié pour décrire la discipline qui étudie les rôles intégrés réciproques des systèmes nerveux et endocrinien dans le contrôle des processus physiologiques.
Avec une compréhension croissante de la neuroendocrinologie, les divisions originales s'effondrent. L'hypothalamus, qui est situé au-dessus et relié à l'hypophyse, est le lien entre les systèmes nerveux et endocrinien, et nombre de ses cellules nerveuses remplissent des fonctions sécrétoires. Il est également lié à d'autres régions majeures du cerveau, notamment le rhinencéphale - le cortex primitif initialement associé à l'olfaction ou à l'odorat - et le système limbique, associé aux émotions. C'est dans l'hypothalamus que sont produites les hormones libérées par l'hypophyse postérieure. L'hypothalamus produit également des substances appelées hormones libératrices et inhibitrices. Ceux-ci agissent sur l'adénohypophyse, l'amenant à augmenter ou à inhiber la production d'hormones de l'hypophyse antérieure, qui agissent sur des glandes situées ailleurs (thyroïde, cortex surrénalien, ovaires, testicules et autres).