Anatomie
L'oreille est l'organe sensoriel responsable de l'audition et du maintien de l'équilibre, via la détection de la position du corps et des mouvements de la tête. Il est composé de trois parties : l'oreille externe, moyenne et interne ; l'oreille externe se trouve à l'extérieur du crâne, tandis que les deux autres parties sont encastrées dans l'os temporal (figure 1).
Figure 1. Schéma de l'oreille.
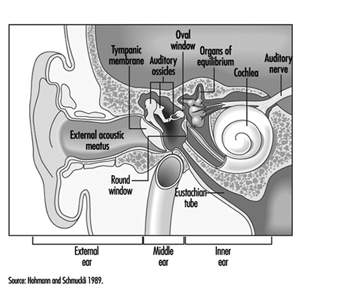
L'oreille externe se compose de l'oreillette, une structure cartilagineuse recouverte de peau, et du conduit auditif externe, un cylindre de forme irrégulière d'environ 25 mm de long qui est tapissé de glandes sécrétant de la cire.
L'oreille moyenne est constituée de la cavité tympanique, une cavité remplie d'air dont les parois externes forment la membrane tympanique (tympan) et communique de manière proximale avec le nasopharynx par les trompes d'Eustache, qui maintiennent l'équilibre de pression de chaque côté de la membrane tympanique. Par exemple, cette communication explique comment la déglutition permet d'égaliser la pression et de restaurer l'acuité auditive perdue causée par un changement rapide de la pression barométrique (par exemple, atterrissage d'avions, ascenseurs rapides). La cavité tympanique contient également les osselets - le marteau, l'enclume et l'étrier - qui sont contrôlés par les muscles stapédien et tenseur du tympan. La membrane tympanique est reliée à l'oreille interne par les osselets, plus précisément par le pied mobile de l'étrier, qui repose contre la fenêtre ovale.
L'oreille interne contient l'appareil sensoriel proprement dit. Il consiste en une coquille osseuse (le labyrinthe osseux) à l'intérieur de laquelle se trouve le labyrinthe membraneux, une série de cavités formant un système fermé rempli d'endolymphe, un liquide riche en potassium. Le labyrinthe membraneux est séparé du labyrinthe osseux par la périlymphe, un liquide riche en sodium.
Le labyrinthe osseux lui-même est composé de deux parties. La partie antérieure est connue sous le nom de cochlée et est le véritable organe de l'ouïe. Il a une forme en spirale rappelant une coquille d'escargot et est pointé vers l'avant. La partie postérieure du labyrinthe osseux contient le vestibule et les canaux semi-circulaires et est responsable de l'équilibre. Les structures neurosensorielles impliquées dans l'audition et l'équilibre sont situées dans le labyrinthe membraneux : l'organe de Corti est situé dans le canal cochléaire, tandis que les macules de l'utricule et du saccule et les ampoules des canaux semi-circulaires sont situées dans la partie postérieure.
Organes auditifs
Le canal cochléaire est un tube triangulaire en spirale, comprenant deux tours et demi, qui sépare la rampe vestibulaire de la rampe tympanique. Une extrémité se termine dans le ligament spiralé, un processus de la colonne centrale de la cochlée, tandis que l'autre est reliée à la paroi osseuse de la cochlée.
La rampe vestibule et le tympan se terminent respectivement par la fenêtre ovale (le pied de l'étrier) et la fenêtre ronde. Les deux chambres communiquent par l'hélicotrème, la pointe de la cochlée. La membrane basilaire forme la surface inférieure du canal cochléaire et supporte l'organe de Corti, responsable de la transduction des stimuli acoustiques. Toutes les informations auditives sont transmises par seulement 15,000 3,500 cellules ciliées (organe de Corti), dont les cellules ciliées dites internes, au nombre de 90 30,000, sont d'une importance critique, car elles forment des synapses avec environ 2 % des XNUMX XNUMX neurones auditifs primaires (figure XNUMX ). Les cellules ciliées internes et externes sont séparées les unes des autres par une abondante couche de cellules de soutien. Traversant une membrane extraordinairement fine, les cils des cellules ciliées sont enchâssés dans la membrane tectoriale, dont l'extrémité libre est située au-dessus des cellules. La surface supérieure du canal cochléaire est formée par la membrane de Reissner.
Figure 2. Coupe transversale d'une anse de la cochlée. Diamètre : environ 1.5 mm.

Les corps des cellules sensorielles cochléaires reposant sur la membrane basilaire sont entourés de terminaisons nerveuses et leurs axones environ 30,000 XNUMX forment le nerf cochléaire. Le nerf cochléaire traverse le conduit auditif interne et s'étend jusqu'aux structures centrales du tronc cérébral, la partie la plus ancienne du cerveau. Les fibres auditives terminent leur chemin tortueux dans le lobe temporal, la partie du cortex cérébral responsable de la perception des stimuli acoustiques.
Organes d'équilibre
Les cellules sensorielles sont situées dans les ampoules des canaux semi-circulaires et les macules de l'utricule et du saccule, et sont stimulées par la pression transmise à travers l'endolymphe à la suite de mouvements de la tête ou du corps. Les cellules se connectent avec des cellules bipolaires dont les processus périphériques forment deux faisceaux, l'un issu des canaux semi-circulaires antérieur et externe, l'autre du canal semi-circulaire postérieur. Ces deux voies pénètrent dans le conduit auditif interne et s'unissent pour former le nerf vestibulaire, qui s'étend jusqu'aux noyaux vestibulaires du tronc cérébral. Les fibres des noyaux vestibulaires s'étendent à leur tour aux centres cérébelleux contrôlant les mouvements oculaires et à la moelle épinière.
L'union des nerfs vestibulaire et cochléaire forme le 8ème nerf crânien, également connu sous le nom de nerf vestibulocochléaire.
Physiologie de l'audition
Conduction du son dans l'air
L'oreille est composée d'un conducteur sonore (l'oreille externe et moyenne) et d'un récepteur sonore (l'oreille interne).
Les ondes sonores traversant le conduit auditif externe frappent la membrane tympanique, la faisant vibrer. Cette vibration est transmise à l'étrier par le marteau et l'enclume. La surface de la membrane tympanique est près de 16 fois celle du pied de l'étrier (55 mm2/3.5mm2), et cela, en combinaison avec le mécanisme à levier des osselets, se traduit par une amplification de 22 fois de la pression acoustique. En raison de la fréquence de résonance de l'oreille moyenne, le rapport de transmission est optimal entre 1,000 2,000 et XNUMX XNUMX Hz. Lorsque le pied de l'étrier bouge, il provoque la formation de vagues dans le liquide à l'intérieur du canal vestibulaire. Le liquide étant incompressible, chaque mouvement vers l'intérieur du pied de l'étrier provoque un mouvement équivalent vers l'extérieur de la fenêtre ronde, vers l'oreille moyenne.
Lorsqu'il est exposé à des niveaux sonores élevés, le muscle de l'étrier se contracte, protégeant l'oreille interne (le réflexe d'atténuation). En plus de cette fonction, les muscles de l'oreille moyenne étendent également la plage dynamique de l'oreille, améliorent la localisation du son, réduisent la résonance dans l'oreille moyenne et contrôlent la pression de l'air dans l'oreille moyenne et la pression du liquide dans l'oreille interne.
Entre 250 et 4,000 80 Hz, le seuil du réflexe d'atténuation est d'environ 0.6 décibels (dB) au-dessus du seuil d'audition, et augmente d'environ 150 dB/dB lorsque l'intensité de stimulation augmente. Sa latence est de 24 ms au seuil, et de 35-10 ms en présence de stimuli intenses. À des fréquences inférieures à la résonance naturelle de l'oreille moyenne, la contraction des muscles de l'oreille moyenne atténue la transmission du son d'environ XNUMX dB. En raison de sa latence, le réflexe d'atténuation offre une protection adéquate contre le bruit généré à des taux supérieurs à deux à trois par seconde, mais pas contre le bruit impulsionnel discret.
La vitesse à laquelle les ondes sonores se propagent dans l'oreille dépend de l'élasticité de la membrane basilaire. L'élasticité augmente, et la vitesse des ondes diminue donc, de la base de la cochlée à la pointe. Le transfert d'énergie vibratoire à la membrane de Reissner et à la membrane basilaire dépend de la fréquence. Aux hautes fréquences, l'amplitude de l'onde est la plus grande à la base, tandis que pour les basses fréquences, elle est la plus grande à la pointe. Ainsi, le point de plus grande excitation mécanique dans la cochlée dépend de la fréquence. Ce phénomène sous-tend la capacité à détecter les différences de fréquence. Le mouvement de la membrane basilaire induit des forces de cisaillement dans les stéréocils des cellules ciliées et déclenche une série d'événements mécaniques, électriques et biochimiques responsables de la transduction mécano-sensorielle et du traitement initial du signal acoustique. Les forces de cisaillement sur les stéréocils provoquent l'ouverture de canaux ioniques dans les membranes cellulaires, modifiant la perméabilité des membranes et permettant l'entrée d'ions potassium dans les cellules. Cet afflux d'ions potassium entraîne une dépolarisation et la génération d'un potentiel d'action.
Les neurotransmetteurs libérés à la jonction synaptique des cellules ciliées internes à la suite de la dépolarisation déclenchent des impulsions neuronales qui voyagent le long des fibres afférentes du nerf auditif vers les centres supérieurs. L'intensité de la stimulation auditive dépend du nombre de potentiels d'action par unité de temps et du nombre de cellules stimulées, tandis que la fréquence perçue du son dépend des populations spécifiques de fibres nerveuses activées. Il existe une cartographie spatiale spécifique entre la fréquence du stimulus sonore et la section du cortex cérébral stimulée.
Les cellules ciliées internes sont des mécanorécepteurs qui transforment les signaux générés en réponse aux vibrations acoustiques en messages électriques envoyés au système nerveux central. Ils ne sont cependant pas responsables du seuil de sensibilité de l'oreille et de son extraordinaire sélectivité en fréquence.
Les cellules ciliées externes, quant à elles, n'envoient aucun signal auditif au cerveau. Au contraire, leur fonction est d'amplifier sélectivement les vibrations mécano-acoustiques à des niveaux proches du seuil par un facteur d'environ 100 (c'est-à-dire 40 dB), et ainsi de faciliter la stimulation des cellules ciliées internes. On pense que cette amplification fonctionne par couplage micromécanique impliquant la membrane tectoriale. Les cellules ciliées externes peuvent produire plus d'énergie qu'elles n'en reçoivent de stimuli externes et, en se contractant activement à des fréquences très élevées, peuvent fonctionner comme des amplificateurs cochléaires.
Dans l'oreille interne, les interférences entre les cellules ciliées externes et internes créent une boucle de rétroaction qui permet de contrôler la réception auditive, en particulier la sensibilité du seuil et la sélectivité en fréquence. Les fibres cochléaires efférentes peuvent ainsi aider à réduire les dommages cochléaires causés par l'exposition à des stimuli acoustiques intenses. Les cellules ciliées externes peuvent également subir une contraction réflexe en présence de stimuli intenses. Le réflexe d'atténuation de l'oreille moyenne, actif principalement aux basses fréquences, et le réflexe de contraction de l'oreille interne, actif aux hautes fréquences, sont donc complémentaires.
Conduction osseuse du son
Les ondes sonores peuvent également être transmises à travers le crâne. Deux mécanismes sont possibles :
Dans le premier, les ondes de compression impactant le crâne provoquent la déformation de la périlymphe incompressible de la fenêtre ronde ou ovale. Comme les deux fenêtres ont des élasticités différentes, le mouvement de l'endolymphe entraîne un mouvement de la membrane basilaire.
Le deuxième mécanisme est basé sur le fait que le mouvement des osselets induit un mouvement uniquement dans la rampe vestibulaire. Dans ce mécanisme, le mouvement de la membrane basilaire résulte du mouvement de translation produit par l'inertie.
La conduction osseuse est normalement inférieure de 30 à 50 dB à la conduction aérienne, comme on le voit facilement lorsque les deux oreilles sont bouchées. Ceci n'est cependant vrai que pour les stimuli à médiation aérienne, la stimulation osseuse directe étant atténuée à un degré différent.
Gamme de sensibilité
Les vibrations mécaniques induisent des changements potentiels dans les cellules de l'oreille interne, les voies de conduction et les centres supérieurs. Seules les fréquences de 16 Hz à 25,000 XNUMX Hz et les pressions acoustiques (elles peuvent être exprimées en Pascal, Pa) de 20 μPa à 20 Pa peut être perçue. La gamme de pressions acoustiques perceptibles est remarquable : une gamme d'un million de fois ! Les seuils de détection de la pression acoustique dépendent de la fréquence, le plus bas à 1 1,000-6,000 XNUMX Hz et augmentant à la fois aux fréquences supérieures et inférieures.
En pratique, le niveau de pression acoustique est exprimé en décibels (dB), échelle de mesure logarithmique correspondant à l'intensité sonore perçue par rapport au seuil auditif. Ainsi, 20 μPa équivaut à 0 dB. Lorsque la pression acoustique décuple, le niveau de décibels augmente de 20 dB, selon la formule suivante :
Lx = 20 bûches Px/P0
où:
Lx = pression acoustique en dB
Px = pression sonore en pascals
P0 = pression acoustique de référence(2×10-5 Pa, le seuil auditif)
Le seuil de discrimination de fréquence, c'est-à-dire la différence de fréquence minimale détectable, est de 1.5 Hz jusqu'à 500 Hz et de 0.3 % de la fréquence du stimulus à des fréquences plus élevées. À des pressions acoustiques proches du seuil auditif, le seuil de discrimination de la pression acoustique est d'environ 20 %, bien que des différences aussi faibles que 2 % puissent être détectées à des pressions acoustiques élevées.
Si deux sons diffèrent en fréquence d'une quantité suffisamment faible, un seul son sera entendu. La fréquence perçue de la tonalité sera à mi-chemin entre les deux tonalités sources, mais son niveau de pression acoustique est variable. Si deux stimuli acoustiques ont des fréquences similaires mais des intensités différentes, un effet de masquage se produit. Si la différence de pression acoustique est suffisamment grande, le masquage sera complet, seul le son le plus fort étant perçu.
La localisation des stimuli acoustiques dépend de la détection du décalage temporel entre l'arrivée du stimulus à chaque oreille et, à ce titre, nécessite une audition bilatérale intacte. Le plus petit décalage temporel détectable est de 3 x 10-5 secondes. La localisation est facilitée par l'effet d'écran de la tête, qui se traduit par des différences d'intensité de stimulus à chaque oreille.
La capacité remarquable des êtres humains à résoudre les stimuli acoustiques est le résultat de la décomposition des fréquences par l'oreille interne et de l'analyse des fréquences par le cerveau. Ce sont les mécanismes qui permettent de détecter et d'identifier des sources sonores individuelles telles que des instruments de musique individuels dans les signaux acoustiques complexes qui composent la musique d'un orchestre symphonique complet.
Physiopathologie
Dommages ciliaires
Le mouvement ciliaire induit par des stimuli acoustiques intenses peut dépasser la résistance mécanique des cils et provoquer une destruction mécanique des cellules ciliées. Comme ces cellules sont en nombre limité et incapables de se régénérer, toute perte cellulaire est permanente, et si l'exposition au stimulus sonore nocif se poursuit, progressive. En général, l'effet ultime des lésions ciliaires est le développement d'un déficit auditif.
Les cellules ciliées externes sont les cellules les plus sensibles aux agents sonores et toxiques tels que l'anoxie, les médicaments ototoxiques et les produits chimiques (par exemple, les dérivés de la quinine, la streptomycine et certains autres antibiotiques, certaines préparations antitumorales), et sont donc les premières à être perdues. Seuls les phénomènes hydromécaniques passifs restent opératoires dans les cellules ciliées externes endommagées ou ayant des stéréocils endommagés. Dans ces conditions, seule une analyse grossière des vibrations acoustiques est possible. En termes très approximatifs, la destruction des cils dans les cellules ciliées externes entraîne une augmentation de 40 dB du seuil auditif.
Dommages cellulaires
L'exposition au bruit, surtout si elle est répétitive ou prolongée, peut également affecter le métabolisme des cellules de l'organe de Corti et des synapses afférentes situées sous les cellules ciliées internes. Les effets extraciliaires rapportés incluent la modification de l'ultrastructure cellulaire (réticulum, mitochondries, lysosomes) et, post-synaptiquement, le gonflement des dendrites afférentes. Le gonflement dendritique est probablement dû à l'accumulation toxique de neurotransmetteurs résultant d'une activité excessive des cellules ciliées internes. Néanmoins, l'étendue des dommages stéréociliaires semble déterminer si la perte auditive est temporaire ou permanente.
Perte auditive due au bruit
Le bruit est un grave danger pour l'ouïe dans les sociétés industrielles de plus en plus complexes d'aujourd'hui. Par exemple, l'exposition au bruit représente environ un tiers des 28 millions de cas de perte auditive aux États-Unis, et le NIOSH (National Institute for Occupational Safety and Health) rapporte que 14 % des travailleurs américains sont exposés à des niveaux sonores potentiellement dangereux. , c'est-à-dire des niveaux supérieurs à 90 dB. L'exposition au bruit est l'exposition professionnelle nocive la plus répandue et la deuxième cause, après les effets liés à l'âge, de perte auditive. Enfin, il ne faut pas oublier la contribution de l'exposition au bruit non professionnel, comme les ateliers à domicile, la musique suramplifiée notamment avec l'utilisation d'écouteurs, l'utilisation d'armes à feu, etc.
Dommages aigus induits par le bruit. Les effets immédiats de l'exposition à des stimuli sonores de haute intensité (par exemple, des explosions) comprennent l'élévation du seuil auditif, la rupture du tympan et des lésions traumatiques de l'oreille moyenne et interne (luxation des osselets, lésion cochléaire ou fistules).
Décalage temporaire du seuil. L'exposition au bruit entraîne une diminution de la sensibilité des cellules sensorielles auditives qui est proportionnelle à la durée et à l'intensité de l'exposition. A ses débuts, cette augmentation du seuil auditif, connue sous le nom de fatigue auditive or décalage de seuil temporaire (TTS), est entièrement réversible mais persiste pendant un certain temps après la fin de l'exposition.
Des études sur la récupération de la sensibilité auditive ont identifié plusieurs types de fatigue auditive. La fatigue à court terme se dissipe en moins de deux minutes et entraîne un décalage maximal du seuil à la fréquence d'exposition. La fatigue à long terme se caractérise par une récupération en plus de deux minutes mais moins de 16 heures, une limite arbitraire dérivée d'études d'exposition au bruit industriel. En général, la fatigue auditive est fonction de l'intensité, de la durée, de la fréquence et de la continuité du stimulus. Ainsi, pour une dose de bruit donnée, obtenue par intégration de l'intensité et de la durée, les modes d'exposition intermittents sont moins nocifs que les modes continus.
La sévérité du TTS augmente d'environ 6 dB pour chaque doublement de l'intensité du stimulus. Au-dessus d'une intensité d'exposition spécifique (le niveau critique), ce taux augmente, en particulier s'il s'agit d'une exposition au bruit impulsif. Le TTS augmente asymptotiquement avec la durée d'exposition ; l'asymptote elle-même augmente avec l'intensité du stimulus. En raison des caractéristiques de la fonction de transfert de l'oreille externe et de l'oreille moyenne, les basses fréquences sont les mieux tolérées.
Les études sur l'exposition aux sons purs indiquent qu'à mesure que l'intensité du stimulus augmente, la fréquence à laquelle le TTS est le plus élevé se déplace progressivement vers des fréquences supérieures à celle du stimulus. Les sujets exposés à un son pur de 2,000 3,000 Hz développent un TTS qui est maximal à environ XNUMX XNUMX Hz (un décalage d'une demi-octave). L'effet du bruit sur les cellules ciliées externes serait responsable de ce phénomène.
Le travailleur qui montre TTS récupère les valeurs auditives de base en quelques heures après avoir été retiré du bruit. Cependant, des expositions répétées au bruit entraînent une moins bonne récupération auditive et une perte auditive permanente qui en résulte.
Décalage de seuil permanent. L'exposition à des stimuli sonores de haute intensité pendant plusieurs années peut entraîner une perte auditive permanente. Ceci est appelé décalage permanent du seuil (PTS). Anatomiquement, le SPT se caractérise par une dégénérescence des cellules ciliées, commençant par de légères modifications histologiques mais aboutissant finalement à une destruction cellulaire complète. La perte d'audition concerne le plus souvent les fréquences auxquelles l'oreille est la plus sensible, car c'est à ces fréquences que la transmission de l'énergie acoustique du milieu extérieur vers l'oreille interne est optimale. Cela explique pourquoi la perte auditive à 4,000 3 Hz est le premier signe de perte auditive d'origine professionnelle (figure XNUMX). Une interaction a été observée entre l'intensité et la durée du stimulus, et les normes internationales supposent que le degré de perte auditive est fonction de l'énergie acoustique totale reçue par l'oreille (dose de bruit).
Figure 3. Audiogramme montrant une perte auditive bilatérale induite par le bruit.
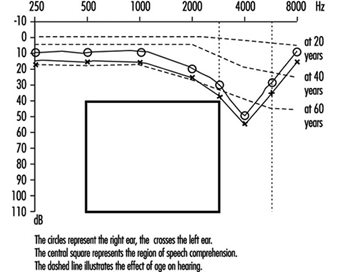
Le développement de la perte auditive induite par le bruit montre une susceptibilité individuelle. Diverses variables potentiellement importantes ont été examinées pour expliquer cette susceptibilité, telles que l'âge, le sexe, la race, les maladies cardiovasculaires, le tabagisme, etc. Les données n'étaient pas concluantes.
Une question intéressante est de savoir si la quantité de TTS pourrait être utilisée pour prédire le risque de SPT. Comme indiqué ci-dessus, il y a un déplacement progressif du TTS vers des fréquences supérieures à celle de la fréquence de stimulation. D'autre part, la plupart des dommages ciliaires survenant à des intensités de stimulation élevées impliquent des cellules sensibles à la fréquence de stimulation. Si l'exposition persiste, la différence entre la fréquence à laquelle le PTS est maximal et la fréquence de stimulation diminue progressivement. Les lésions ciliaires et la perte cellulaire se produisent par conséquent dans les cellules les plus sensibles aux fréquences de stimulation. Il apparaît donc que TTS et PTS impliquent des mécanismes différents, et qu'il est donc impossible de prédire le PTS d'un individu sur la base du TTS observé.
Les personnes atteintes du SPT sont généralement asymptomatiques au départ. Au fur et à mesure que la perte auditive progresse, ils commencent à avoir des difficultés à suivre les conversations dans des environnements bruyants tels que des fêtes ou des restaurants. La progression, qui affecte généralement la capacité à percevoir les sons aigus en premier, est généralement indolore et relativement lente.
Examen des personnes souffrant de perte auditive
Examen clinique
En plus de l'historique de la date à laquelle la perte auditive a été détectée pour la première fois (le cas échéant) et de son évolution, y compris toute asymétrie de l'audition, le questionnaire médical doit obtenir des informations sur l'âge du patient, ses antécédents familiaux, l'utilisation de médicaments ototoxiques ou l'exposition à d'autres produits chimiques ototoxiques, la présence d'acouphènes (c'est-à-dire des bourdonnements, des sifflements ou des bourdonnements dans une ou les deux oreilles), des étourdissements ou tout problème d'équilibre, et tout antécédent d'infection de l'oreille avec douleur ou écoulement du conduit auditif externe. Il est d'une importance cruciale de disposer d'un historique détaillé des expositions à des sonner niveaux (notez que, pour le profane, tous les sons ne sont pas du « bruit ») au travail, dans les emplois précédents et en dehors du travail. Un historique des épisodes de STT confirmerait les expositions toxiques antérieures au bruit.
L'examen physique doit inclure une évaluation de la fonction des autres nerfs crâniens, des tests d'équilibre et une ophtalmoscopie pour détecter tout signe d'augmentation de la pression crânienne. L'examen visuel du conduit auditif externe détectera tout cérumen impacté et, après l'avoir retiré avec précaution (pas d'objet pointu !), tout signe de cicatrisation ou de perforation de la membrane tympanique. La perte auditive peut être déterminée très grossièrement en testant la capacité du patient à répéter des mots et des phrases prononcés à voix basse ou chuchotés par l'examinateur lorsqu'il est placé derrière et hors de la vue du patient. Le test de Weber (placement d'un diapason vibrant au centre du front pour déterminer si ce son est "entendu" dans l'une ou les deux oreilles) et le test du diapason de Rinné (placement d'un diapason vibrant sur l'apophyse mastoïdienne jusqu'à ce que le patient ne peut plus entendre le son, puis placer rapidement la fourche près du conduit auditif ; normalement, le son peut être entendu plus longtemps dans l'air que dans les os) permettra de classer la perte auditive comme transmission ou neurosensorielle.
L'audiogramme est le test standard pour détecter et évaluer la perte auditive (voir ci-dessous). Des études spécialisées pour compléter l'audiogramme peuvent être nécessaires chez certains patients. Il s'agit notamment de : tympanométrie, tests de discrimination des mots, évaluation du réflexe d'atténuation, études électrophysiques (électrocochléogramme, potentiels évoqués auditifs) et radiologiques (radiographies crâniennes de routine complétées par CAT scan, IRM).
Audiométrie
Cet élément crucial de l'évaluation médicale utilise un appareil appelé audiomètre pour déterminer le seuil auditif des individus à des sons purs de 250-8,000 10 Hz et des niveaux sonores entre -110 dB (seuil auditif des oreilles intactes) et 16 dB (dommages maximaux ). Pour éliminer les effets des TTS, les patients ne doivent pas avoir été exposés au bruit au cours des 3 heures précédentes. La conduction aérienne est mesurée par des écouteurs placés sur les oreilles, tandis que la conduction osseuse est mesurée en plaçant un vibreur en contact avec le crâne derrière l'oreille. L'audition de chaque oreille est mesurée séparément et les résultats des tests sont rapportés sur un graphique appelé audiogramme (Figure XNUMX). Le seuil d'intelligibilité, c'est-à-dire. l'intensité sonore à laquelle la parole devient intelligible, est déterminée par une méthode de test complémentaire appelée audiométrie vocale, basée sur la capacité à comprendre des mots composés de deux syllabes d'intensité égale (par exemple, berger, dîner, étourdir).
La comparaison de la conduction aérienne et osseuse permet de classer les pertes auditives en perte de transmission (impliquant le conduit auditif externe ou l'oreille moyenne) ou neurosensorielle (impliquant l'oreille interne ou le nerf auditif) (figures 3 et 4). L'audiogramme observé en cas de surdité due au bruit se caractérise par un début de surdité à 4,000 3 Hz, visible par un creux de l'audiogramme (figure 3,000). Au fur et à mesure que l'exposition à des niveaux de bruit excessifs se poursuit, les fréquences voisines sont progressivement affectées et le creux s'élargit, empiétant, à environ 15 500 Hz, sur des fréquences essentielles à la compréhension de la conversation. La perte auditive induite par le bruit est généralement bilatérale et présente un schéma similaire dans les deux oreilles, c'est-à-dire que la différence entre les deux oreilles ne dépasse pas 1,000 dB à 2,000 Hz, à 30 3,000 dB et à 4,000 6,000 Hz, et 4,000 dB à 4 XNUMX, à XNUMX XNUMX et à XNUMX XNUMX Hz. Des lésions asymétriques peuvent cependant être présentes en cas d'exposition non uniforme, par exemple chez les tireurs d'élite, chez qui la perte auditive est plus élevée du côté opposé au doigt à ressaut (côté gauche, chez un droitier). Dans les surdités non liées à l'exposition au bruit, l'audiogramme ne présente pas le creux caractéristique de XNUMX XNUMX Hz (figure XNUMX).
Figure 4. Exemples d'audiogrammes de l'oreille droite. Les cercles représentent la perte auditive par conduction aérienne, la « conduction osseuse ».
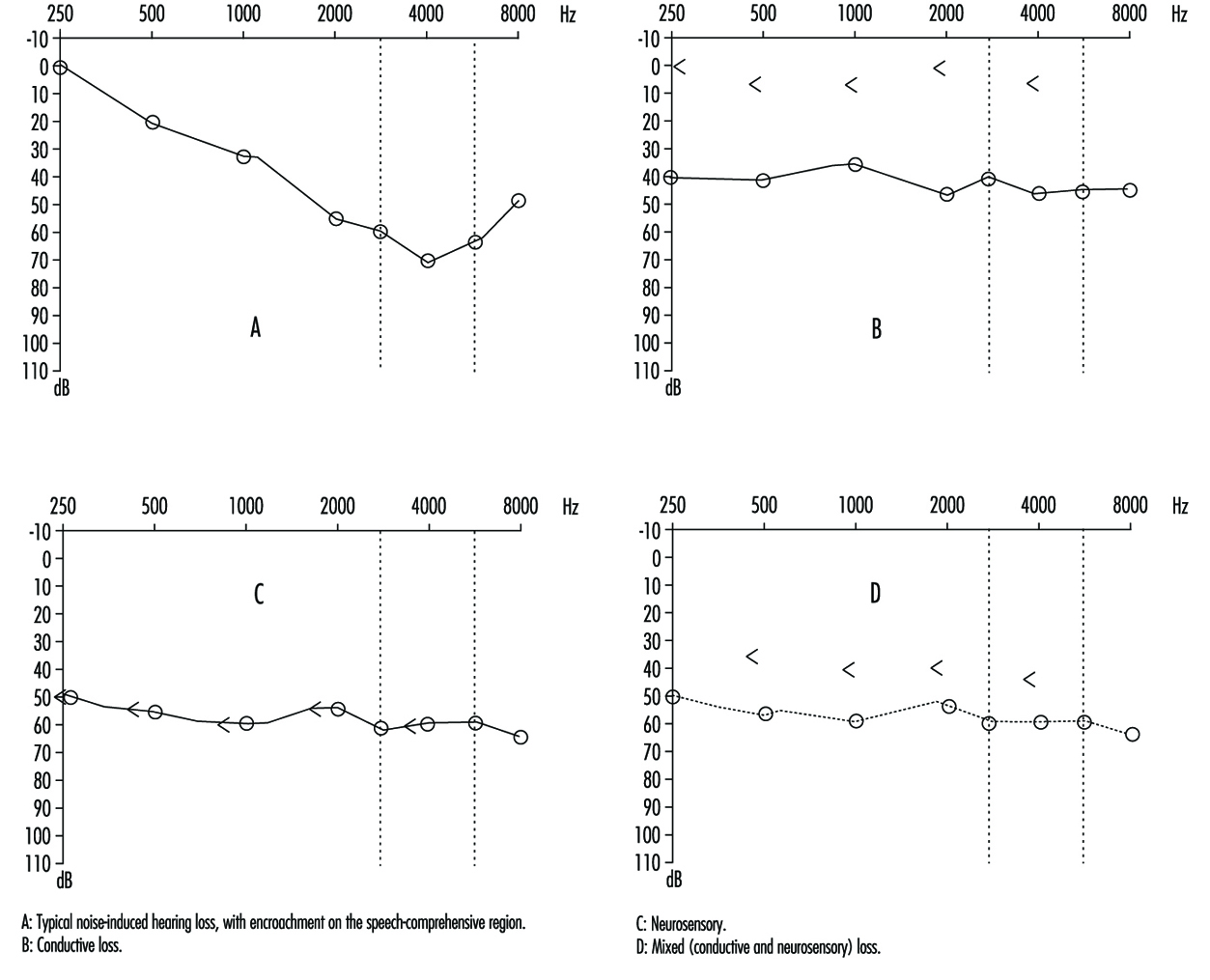
Il existe deux types d'examens audiométriques : de dépistage et de diagnostic. L'audiométrie de dépistage est utilisée pour l'examen rapide de groupes d'individus sur le lieu de travail, dans les écoles ou ailleurs dans la communauté afin d'identifier ceux qui apparaître avoir une certaine perte auditive. Souvent, des audiomètres électroniques qui permettent l'autotest sont utilisés et, en règle générale, les audiogrammes de dépistage sont obtenus dans un endroit calme, mais pas nécessairement dans une chambre insonorisée et exempte de vibrations. Cette dernière est considérée comme une condition préalable à l'audiométrie diagnostique qui vise à mesurer la perte auditive avec une précision et une exactitude reproductibles. L'examen diagnostique est correctement effectué par un audiologiste qualifié (dans certaines circonstances, une certification formelle de la compétence de l'audiologiste est requise). La précision des deux types d'audiométrie dépend des tests périodiques et du recalibrage de l'équipement utilisé.
Dans de nombreuses juridictions, les personnes atteintes d'une perte auditive liée au travail et due au bruit sont éligibles aux prestations d'indemnisation des accidents du travail. En conséquence, de nombreux employeurs incluent l'audiométrie dans leurs examens médicaux préalables à l'embauche pour détecter toute perte auditive existante qui pourrait être la responsabilité d'un employeur précédent ou représenter une exposition non professionnelle.
Les seuils d'audition augmentent progressivement avec l'âge, les fréquences plus élevées étant plus touchées (figure 3). La baisse caractéristique de 4,000 XNUMX Hz observée dans la perte auditive due au bruit n'est pas observée avec ce type de perte auditive.
Calcul de la perte auditive
Aux États-Unis, la formule la plus largement acceptée pour calculer la limitation fonctionnelle liée à la perte auditive est celle proposée en 1979 par l'American Academy of Otolaryngology (AAO) et adoptée par l'American Medical Association. Elle est basée sur la moyenne des valeurs obtenues à 500, à 1,000 2,000, à 3,000 1 et à 25 XNUMX Hz (tableau XNUMX), la limite inférieure de limitation fonctionnelle étant fixée à XNUMX dB.
Tableau 1. Calcul typique de la perte fonctionnelle à partir d'un audiogramme
| La fréquence | |||||||
| 500 Hz |
1,000 Hz |
2,000 Hz |
3,000 Hz |
4,000 Hz |
6,000 Hz |
8,000 Hz |
|
| Oreille droite (dB) | 25 | 35 | 35 | 45 | 50 | 60 | 45 |
| Oreille gauche (dB) | 25 | 35 | 40 | 50 | 60 | 70 | 50 |
| Perte unilatérale |
| Pourcentage de perte unilatérale = (moyenne à 500, 1,000 2,000, 3,000 XNUMX et XNUMX XNUMX Hz) – 25dB (limite inférieure) x1.5 |
| Mise en situation : Oreille droite : [([25 + 35 + 35 + 45]/4) – 25) x 1.5 = 15 (pour cent) Oreille gauche : [([25 + 35 + 40 + 50]/4) – 25) x 1.5 = 18.8 (pour cent) |
| Perte bilatérale |
| Pourcentage de perte bilatérale = {(pourcentage de perte unilatérale de la meilleure oreille x 5) + (pourcentage de perte unilatérale de la pire oreille)}/6 |
| Exemple: {(15 x 5) + 18.8}/6 = 15.6 (pour cent) |
Source : Rees et Duckert 1994.
Presbyacousie
La presbyacousie ou perte auditive liée à l'âge commence généralement vers l'âge de 40 ans et progresse progressivement avec l'âge. Elle est généralement bilatérale. La baisse caractéristique de 4,000 XNUMX Hz observée dans la perte auditive due au bruit n'est pas observée avec la presbyacousie. Cependant, il est possible que les effets du vieillissement se superposent à la perte auditive liée au bruit.
Traitement
Le premier élément essentiel du traitement est d'éviter toute nouvelle exposition à des niveaux de bruit potentiellement toxiques (voir « Prévention » ci-dessous). On pense généralement qu'il ne se produit pas plus de perte auditive ultérieure après la suppression de l'exposition au bruit que ce à quoi on pourrait s'attendre du processus de vieillissement normal.
Alors que les pertes de conduction, par exemple, celles liées à des dommages aigus induits par le bruit traumatique, peuvent faire l'objet d'un traitement médical ou d'une intervention chirurgicale, la perte auditive chronique induite par le bruit ne peut pas être corrigée par un traitement. L'utilisation d'une prothèse auditive est le seul « remède » possible, et n'est indiquée que lorsque la perte auditive affecte les fréquences critiques pour la compréhension de la parole (500 à 3,000 XNUMX Hz). D'autres types de support, par exemple la lecture labiale et les amplificateurs de son (sur les téléphones par exemple), peuvent cependant être possibles.
Prévention
La surdité due au bruit étant permanente, il est indispensable d'appliquer toute mesure susceptible de réduire l'exposition. Cela inclut la réduction à la source (machines et équipements plus silencieux ou enfermés dans des enceintes insonorisées) ou l'utilisation de dispositifs de protection individuelle tels que des bouchons d'oreille et/ou des casques antibruit. Si l'on se fie à ces derniers, il est impératif de vérifier que les revendications d'efficacité de leurs fabricants sont valables et que les travailleurs exposés les utilisent correctement à tout moment.
La désignation de 85 dB (A) comme limite d'exposition professionnelle la plus élevée autorisée visait à protéger le plus grand nombre de personnes. Mais, comme il existe des variations interpersonnelles importantes, des efforts acharnés pour maintenir les expositions bien en deçà de ce niveau sont indiqués. Une audiométrie périodique doit être mise en place dans le cadre du programme de surveillance médicale afin de détecter le plus tôt possible tout effet pouvant indiquer une toxicité sonore.